第十章 叶足虫
第一节溶组织内阿米巴
第二节其他消化道阿米巴
一、哈门氏内阿米巴
二、结肠内阿米巴
三、微小内蜒阿米巴
四、布氏嗜碘阿米巴
五、齿龈内阿米巴
第三节致病性自生生活阿米巴
叶足虫隶属于肉足鞭毛门(Phylum Sarcomastigophora)的叶足纲(Class Lobosea),以具有宽大叶状伪足的细胞运动器为基本特征。多数种的生活型含一个形态各异的泡状核,营无性繁殖,一般有滋养体和包囊两个生活史期,个别种缺包囊期。寄生人体的常见种类多为消化道腔道型原虫,计有3属7种(见表9-1),均归属于内阿米巴科(Family Entamoebidae),其中仅溶组织内阿米巴一个种对人致病,可引起侵袭型阿米巴病(invasive amoebiasis)。少数自生生活类型的非内阿米巴科的种类亦可偶然侵入人体引起严重疾病。叶足纲原虫科以上的分类归属尚有不少争议。
第一节 溶组织内阿米巴
溶组织内阿米巴(Entamoeba histolytica Schaudinn,1903),即痢疾阿米巴,为侵袭型阿米巴病的病原虫,主要寄生于结肠,引起阿米巴痢疾和各种类型的阿米巴病,为全球分布,多见于热带与亚热带。据统计,在全球超过5亿的阿米巴感染者中,侵袭型的年发病率高达4千万例以上,至今每年死于阿米巴病的人数不少于4万,当前在医学上的重要性已被认为仅次于疟疾与血吸虫病。
人类对阿米巴病早有认识与记载。古希腊希波克拉蒂对痢疾的潜在传染性和并发症-肝泻(hepatic flux)已有明确论述;我国古代医书《内经素问》《伤寒论》等也记有“下痢”,“赤痢”,“疫痢”等有关鉴别诊断的临床经验。阿米巴痢疾的病原体则由俄罗斯医师 лйщ于1975年首先发现,至1903年由德国著名微生物学家Schaudinn定名。对于溶组织内阿米巴的生物学、种群毒力及其致病作用和机制的研究已历经100多年,直到20世纪60年代末美国学者Diamond开拓无共生物纯培养之后,才为细胞及分子水平的生物、生化及免疫学研究提供了高纯高的虫体材料,从而取得新的进展。虽然还留有不少有待深入探讨的问题,对于持续了大半个世纪有关种株与毒力的争论热点,已有初步结论,从生化构成、免疫原性以及基因分析确认了英国学者Brumpt早在1925年即已提出的善于致病与非致病品系属于不同种群的论断。
形态与生活史
近代研究表明,溶组织内阿米巴应是一个包含两种在形态学难于区分而致病力显著不同的种群复合体,即存在着侵袭型和非侵袭型(或称致病和非致病型或弱致病型)两个不同的种。长期以来,人们试图以此解释阿米巴感染的低发病率(<10%)现象,但只有通过70年代末期以来从生化、免疫以及遗传研究的不断探索,终于获得了细胞与分子生物学的确凿证据,并根据研究提供的信息的数据,Diamond和Clark(1993)确认这两个种的存在,重新描述了沿用Entamoeba histolytica Schaudinn,1903学名的侵袭型种的形态学,用以区分非致病种,Entamoeba dispar Brumpt,1925。鉴于这两个种的区分目前仅借助于生化与基因分析手段,对于溶组织内阿米巴的有关生物学描述,仍须借鉴传统的复合群体概念。
溶组织内阿米巴的生活史有滋养体、囊前期、包囊和囊后滋养体各期(图10-1,2),其中囊前期和囊后滋养体分别是滋养体转化为包囊和包圳转化为滋养体的短暂过渡期,仅出现于肠道寄生阶段。整个生活史过程仅须一种哺乳类宿主,人是主要的适宜宿主。猿、猕猴、犬、猪、鼠均有自然或实验感染的报道,但并无重要流行病意义。

图10-1 溶组织内阿米巴 (铁苏木素染色)
滋养体期系该阿米巴的基本生活型,通常在结肠腔内以二分裂法繁殖。直径10~40µm,个体大小随种系而有差异,可大至60µm以上。在光镜下观察活体,可见较白细胞稍大的折光性活动小体,在适宜温度下运动活泼,常伸出单一伪足作定向阿米巴运动。扫描电镜观察,可见在表膜的一个部位附有许多小泡,为伪尾区,可能是一个排废部位。滋养体需经固定染色后方可辨明内部结构。最常用铁苏木素染色,在高倍放大的光镜下,胞质可辨认较透明的外质和颗粒状内质,内质含一典型泡状核,直径4~7µm,在不着色的纤薄核膜内缘有排列整齐的单层染色质粒(chromatin granules),有一个位于正中或稍偏位的粒状核仁,核仁与核膜之间隐约可见网状核纤丝。典型的核结构有助于虫种鉴别;胞质内含食物泡及吞噬的红细胞。电镜下,滋养体的外被可见由两层电子致密层组成的典型单位膜结构及外层绒毛状糖萼外被。后者的厚度在肠壁病灶分离的滋养体常数倍于无共生物培养的个体。表膜还分布有许多含肌动蛋白的丝状突起为丝状伪足,可证明参与阿米巴的侵袭机制。扫描电镜下的表膜还可见大小不一的杯状吞噬微口(phagocytic stomata),司吞噬与吞饮的功能。内质中可见众多的食物泡、空泡和大小不等的糖原粒,另有微管、溶酶体及小泡状或管状内质网,迄今未发现有线粒体、糙面内质网和典型的高尔基复合体。
在肠腔内增殖的滋养体可随肠内容物下移,随着肠内环境的变化,如水分逐渐被吸收等,停止活动,团缩,排出未消化食物,形成囊前期,此期胞质内有时可出现一种特殊的营养储存结构 ──拟染色体(chromatoid body),经分析系由90%以上的核糖核蛋白体聚合而成,能被深染,呈现独特的短棒状,为虫种鉴别的形态学特点。囊前期形成后,胞质分泌囊壁包裹于质膜之外,形成圆形的包囊期。包囊直径在10~20µm之间,电镜下可见囊壁为双层。包囊初期只具一个胞核,随后二次分裂为4核,偶见8核,粪便中可查到成熟度不同的1核、2核或成熟的4核包囊,核的结构同滋养体期,在1或2核包囊可见糖原泡和棍状的拟染色体。碘染时包囊呈淡棕色或黄色,糖原泡为棕红色(彩图Ⅲ)铁苏木素染色的包囊为深蓝色,糖原泡被溶解成空泡,拟染色体色深更清晰。包囊为阿米巴的传播阶段,仅见于宿主的粪便内,4核包囊具感染性,在传播上起重要作用,为阿米巴的传播阶段,仅见于宿主的粪便内,4核包囊具感染性,在传播上起重要要作用,入侵组织的滋养体不形成包囊,复合体内侵袭与非侵袭种的包囊则无法从形态学上加以肠下段经硷性消化液作用后,囊壁变薄,虫体活跃,随即脱囊而出形成含4个胞核的囊后滋养体。此期历时甚短,脱囊后核很快各分裂一次,继之胞体分为8个个体较小的小滋养体,在回盲部定居于结肠粘膜皱褶或肠腺窝间,以宿主肠粘液、细菌及已消化食物为营养,发育至一定大小后不断以二分裂增殖。在肠腔内的滋养体称小滋养体,虫体较小(10~20µm)也称肠腔共栖型。小滋养体在某种因素影响下可不同程度地入侵肠壁 ,可吞噬红细胞和组织细胞转变为大滋养体,也称组织型滋养体,其体积较大(20~40µm),运动活泼,并不断破坏肠壁组织,引起原发病灶;侵入肠组织的滋养体可随血流至肝或其他部位;也可能随坏死组织脱落入肠腔,在急性期肠蠕动增加的情况下,大滋养体无成囊的充分时间,致使大量排出体外。(图10-2)。由于滋养体对外环境的抵御力很弱,故在传播上不起作用。而在无症状带虫者的正常粪便中可排出大量包囊,成为流行病学上的重要传染源。据估计一个带虫者每天排出的包囊可逾5000多个。
生理代谢
溶组织内阿米巴营兼性厌氧代谢,主要以酵解糖原获取能量。在代谢通路上因缺细胞色素及经典三羧酸循环等酶系而拥有数种特有酶系参加糖的分解反应,例如电子转运蛋白相关的还原酶系复合物可使虫体在有氧环境下有限度地利用氧,人们已利用这一酶系的还原作用研究出对阿米巴具有选择毒性作用的5-硝基咪唑类药物;溶组织内阿黧米巴可以补救途径利用各种嘌呤、嘧啶和相应的核苷合成RNA和DNA。有选择地阻断阿米巴特有的酶系作用或DNA,RNA的合成途径可用为筛选特效化疗药物的有效手段。
溶组织内阿米巴体有大量多糖贮存,在培养条件下它们能特异地利用某些糖类。实验证明米淀粉能提供滋养体最佳生长的营养条件,所以常规培养基内必须加入适量米粉,以及血清、类酯(胆固醇等)、无机盐类、多种维生素等。在肠腔或人工培养基上,阿米巴原虫与某些细菌有明显的共生现象,共生菌不仅是肠腔内阿米巴的食料,还可造成对原虫有利的相对厌氧环境,并维持适度的酸碱条件(pH)。对溶组织内阿米巴营养需求和生存条件的研究已为其体外人工培养提出不断改进,从30年代的有菌培养(xenic culture),到50年代的单栖培养(monoxenic culture),直至60年代的无共生物纯培养(axenic culture)及70年代的软琼脂克隆培养作出了贡献。

图10-2溶组织内阿米巴生活史
溶组织内阿米巴对宿主组织的侵袭力特异地表现为一种对靶物的接触性杀伤功能。这种“触杀”机制,是一个包括对靶细胞和组织的粘附、杀伤、溶噬的连锁生理过程。滋养体表膜已现有两种具植物凝血素(lectin)活性的粘附因子(adhesin)能与靶细胞糖萼中的乙酰氨基葡萄糖(GLcNAc)和乙酰氨基兰乳糖胺(GalNAc)发生受体性结合,提供了对靶细胞进行攻击的前提。已证明此种粘附活性与虫种毒力相关。阿米巴的杀伤作用是由酶溶、细胞毒、胞噬以及协同损伤等构成的综合机制。溶组织内阿米巴原虫在完成对宿主细胞的粘附、胞溶以后即随之以胞噬,而吞噬的活力则亦与虫株的的毒力和胞溶力一致。实验也证明阿米巴对宿主中性粒细胞有化学趋附性。粒细胞被滋养体触溶后释出有毒氧化物而加剧组织破坏,说明溶组织内阿米巴的侵袭并存着宿主细胞的协同作用。
致病机制
溶组织内阿米巴的致病作用是虫体和宿主的相互作用,并受多种因素影响的复杂过程。人被感染后可呈从无疾病带虫状态到急性痢疾或脓肿的各种临床类型,病理和病程复杂多变。阿米巴病的发生发展与原虫的毒力、寄生微环境的理化、生物因素以及宿主机体状态等有密切的关系。
善于种株毒力问题已如前述。根据近年来流行病学研究,同工酶分析,特异单克隆抗体测试,基因克隆的印证,致病与非致病种群的客观存在已无疑议。近十年来,全世界各地分离出6000多株溶组织内阿米巴分离株,发现了22种同工酶谱型(zymodeme),据此,可分为与临床致病完全相一致的侵袭与非侵袭型两类谱型;随后,分别制备的单克隆抗体也相应展示了两者的抗原特异性;而近年进行的基因指纹、核酸杂交以及结合多聚酶链反应的DNA扩增技术进一步显示了它们在遗传属性上是两个种群。现在,已可借助多种技术对分离株或样本中的病原进行种群鉴定。但是,阿米巴感染的发病还受虫株毒力以外的众多因素影响,其中宿主肠道内环境,特别是共生菌群的作用十分显著。志愿者实验表明吞食无菌包囊只发生带虫状态,随后吞服病人肠道菌种就发生了痢疾;国内外动物实验也证明,用混加产气荚膜杆菌等多种细菌的阿米巴感染小猫或豚鼠,其感染率均较无菌对照组为高,所致病变也较重。此种促成致病协同作用的菌群除提供有利于阿米巴增殖活动的理化条件外,还可能削弱宿主的全身或局部抵抗力,甚至直接损害肠粘膜,为阿米巴侵入组织提供条件。另外,疾病阿米巴必须突破宿主的防卫屏障才能侵入组织繁殖。
由囊后滋养体增殖的虫体群大多在回盲部或少数在乙状结肠引起原发病灶。除重症者外,原发灶仅局限于粘膜层,肠窥镜观察为稍隆起的充血小灶,中央常有针尖状溃破口。无细菌伴发感染时周围无明显炎症反应。在急性病例,增殖的滋养体能突破粘膜肌层,在疏松的粘膜下层繁殖扩展,引起液化坏死灶,形成口小底大的烧瓶样溃疡。在溃疡处可查见滋养体(图10-3)。溃疡间的粘膜常正常,甚至与机体的修复机制并存,这与菌痢引起的弥漫性病灶迥然不同。继发溃疡的好发部位以回盲部及乙状结肠最多,结肠的弯曲部分次之。病大小自数毫米至十多毫米不等,若有细菌伴发感染,则镜检可见坏死灶的外围有多量中性粒细胞浸润。重症病例则溃疡可深入肌层,或邻近的溃疡互相融合,致使大片粘膜脱落,因而阿米巴溃疡具有较大穿孔肌层,轻症或亚急性病例的肠壁损害可有粘膜微小溃疡至肉芽肿样溃疡的各种不同类型。慢性病例由于粘膜增生可出现阿米巴肿(amoeboma)。在肠粘膜下层或肌层的阿米巴滋养体可侵入静脉,随血流截留于肝窦,引起继发性阿米巴病。初起为多发性坏死小灶,与病灶外围的白细胞浸润构成肝炎期。根据宿主的机体状况,其中的一个或偶尔多个小灶融合而发展为肝脓肿。脓肿的中央为含酱色性状的坏死区,滋养体则聚集在炎症病灶外围与正常组织交界的部位(脓肿壁)。大的肝脓肿可发展至婴儿头颅大小。此外痢疾阿米巴也可经血路或直接经膈向胸腔穿破入肺而致肺脓肿;侵入纵隔、心包甚至脑、脾等部位引起局部脓肿,但甚为少见。肝脓肿腹壁穿孔部位,手术切口或会阴附近的皮肤也可被侵袭而发生阿米巴皮肤溃疡;如累及生殖器官则可引起阿米巴性阴道炎或前裂腺炎等。

图10-3阿米巴痢疾肠壁溃疡切片
1.示口小底大型溃疡; 2.示溃疡组织内阿米巴滋养体( ↑)
(采 Markell等)
阿米巴病的临床表现较多变化,常有迁延现象,即病程延长,症状隐显无常。按WHO建议的临床分型可分为无症状的带虫感染和有症状的侵袭性感染。前者占90%以上,绝大多数系复合体中非侵袭种的感染。后者又分肠阿炎巴病(包括阿米巴痢疾、肠炎,阿米巴肿,阿米巴性阑尾炎等)和肠外阿米巴病(包括阿米巴肝、肺、脑脓肿及皮肤阿米巴病等)两类。典型的阿米巴痢疾以常伴有腹绞痛及里急后重、含脓血糖液便的急性腹型的痢疾已不多见,大多表现为亚急性或慢性迁延性肠炎,可伴有腹胀、消瘦、贫血等。肠外阿米巴病以阿米巴肝脓肿最多见,系血行播散,好发于肝右叶,常伴肠阿米巴病史,大多起病缓慢,有弛张热、肝肿大、肝区痛及进行性消瘦、贫血和营养性水肿等。阿米巴肺脓肿较少见,有肝源性和肠源性。前者多由阿米巴肝脓肿直接穿破所致;后 纱血路传播,病灶不限于右下叶。极少数情况下,肝脓肿可穿入心包、穿破腹壁。肠道阿米巴也可进入肛周、阴道、尿道等引起相应部位的脓肿或炎症。
宿主感染溶组织内阿米巴后可激发体液及细胞免疫应答。应用各种免疫学方法如IHA和ELISA等均可测到阿米巴病患者血中的特异抗体,除各IgG亚类外也有IgM,IgA和IgE。IgE抗体在治愈一年内滴度虽明显跌降,但仍维持高于正常值数倍达10年以上,而IgM和IgE则多数于治愈90天内回复至原有水平。若病后3个月IgM和IgE才出现或重新出现,提示预后不良。除血清抗体以外,约80%的阿米巴病患者还可测到以IgA为主体的粪抗体(copro-antibodies)。实验研究表明,抗阿米巴抗体的水平与保护作用并不一致,而感染所激发的细胞免疫在抗虫机制中起重要作用。已证明侵袭型阿米巴的虫体可溶性抗原可以活化从阿米巴病人分离的淋巴细胞,分泌含丰富 γ-干扰素的细胞因子,进而活化巨噬细胞和中性粒细胞,使增强吞噬力,产生超氧化物,杀伤滋养体,在体外还可抑制阿米巴蛋白质及DNA的合成。但是,阿米巴病的早期,细胞免疫多呈抑制状态,到后期才转化为有免疫保护性。这是因为阿米巴的抗原组成中含有激发免疫抑制的决定簇,在疫苗研制中应引起特别注意。此外,生活的溶组织内阿米巴具有独特的逃避宿主免疫攻击的机制,例如致病型阿米巴滋养体对补体介导的溶解作用具抵抗力;滋养体还可利用质膜能运动的特点把结合于虫体表面的抗体,不断地被一种“帽化”(capping)机制(即把复合物集中并向后移动),通过uroid区周期性地由体表脱落而被清除。
实验诊断
1.病原检查
确诊肠阿米巴病的方法,常用的有粪便检查、人工培养和肠镜活组织检查或刮拭物涂片检查。
⑴粪便检查:
1)生理盐水涂片法:适用于急性痢疾患者的脓血便或阿米巴肠炎的稀便,主要检查活动的滋养体,但标本必须新鲜,送检越快越好,置4℃不宜超过4~5小时。典型的阿米巴痢疾粪便为酱红色粘液样,有腥臭味。镜检可见粘液里含很多粘集成团的红细胞和较少的白细胞,有时可见棱形结晶(Charcot-Leyden crystal)和活动的滋养体。这些特点可与细菌性痢疾的粪便区别。
2)包囊浓集法:对慢性患者的成形粪样,亦可用直接涂片法查找包囊期,常作碘液染色以显示细胞核,便于鉴别诊断。但包囊检查可用浓集法提高检出率。常用的方法有硫酸锌浮集聚法和汞碘醛离心沉淀法(MIFC,详附录)。
临床上多见的不典型迁延型阿米巴病,常不易在粪便内找到病原体。据分析,无症状患者或病变局限于盲肠和升结肠者,常规湿涂片或固定染色涂片的一次检出率不超过30%,间隔一天以上的3次送检,阳性率可提高至60~80%,送5次者可达90%以上。
⑵人工培养:已有多种改良的培养基可供选用(见附录)。从粪便标本分离培养虫体的诊断常规都用有菌培养,但一般在多数亚急性或慢性病例的检出率不高,故培养法似不宜用作常规检查。无共生物培养需特种培养基和技术要求,宜用于研究。
⑶组织检查:借助乙状结肠镜或纤维结肠镜直接观察粘膜溃疡并作活检或刮拭物涂片的检出率最高,约85%的痢疾患者可用此法检出。活体标本必须取材于溃疡边缘,脓腔穿刺亦应取材于壁部,并注意脓液性状特征。
病原检查要特别注意盛器洁净及病人服药和治疗措施的影响。某些抗生素,杀虫药物,泻剂、收敛剂,高、低渗灌肠液,钡餐以及自身尿液污染均可使滋养体致死而干扰病原体的检出。
2.免疫诊断 由于阿米巴病的病原查诊容易漏检与误诊,免疫学诊断虽属间接的辅诊手段,却具有很大的实用价值。自60年代阿米巴无共生物培养建立和特异单克隆抗体问世后,提供了溶组织内阿米巴纯抗原和优质工具抗体,国内外陆续发展了多种免疫诊断方法。近年以酶联免疫吸附试验(ELISA)的各种改良法应用较多。大体上,特异循环抗体的检测在肝脓肿患者的检出率可高达95%~100%,侵袭型肠病患者85%~95%,而无症状带虫者仅10%~40%,滴度视病情可不一致,但脓肿大者多为高滴度。因此血清学诊断仅对急性发病患者有较大的辅助诊断价值,血清流行病学调查中,人群抗体滴度的消长水平可提示地区发病情况。单克隆抗体和DNA探针杂交技术的应用为探测宿主血液和排泄物中的病原物质提供了特异、灵敏并能抗干扰的示踪工具。应用单克隆抗体检测粪便、脓液内虫源抗原及DNA探针鉴定粪内虫种均已见有报告。
流行病学
阿米巴病呈世界性分布,全球高发地区位于墨西哥、南美洲东部、东南亚,西非等,地处北纬10°至南纬10°之间的热带和亚热带地区,平均感染率在20%以上,个别地区如埃及可达57%~87%。据报告,我国近年的人群感染率在0.7%~2.17%之间,大多见于经济条件、卫生状况、生活环境较差的地区,农村高于城市。随着改革形势的推进,我国全民的生活卫生水平有所提高,除个别边远地区外,典型的急性阿米巴病例已属少见,多数为散在分布的迁延型患者或带虫者。目前,我国的肠阿米巴病发病率低于肝脓肿的发病率。在欧美则高发人群主要为男性同性恋者和旅游者。在全球近5亿感染者中,侵袭型的发病患者约占10%,肠阿米巴病的发病率为阿米巴肝脓肿的5~50倍,后者的病死率为2%~10%,爆发性阿米巴肠炎则高达70%,估计每年死亡人数仅次于疟疾和血吸虫病,列为世界上死于寄生虫病的第三位。
阿米巴病的传染源主要为粪便里持续有包囊排出的带虫者。包囊在外环境具较强的生存力,在潮湿低温环境可存活12天以上,水中可活9~30天,但以干燥、高温和化学药品的抗力不强,却可无损伤地通过蝇或蟑螂的消化道。阿米巴滋养体在体外极易死亡,无传播作用。溶组织内阿米巴没有重要的保虫宿主,人群的感染主要通过人际经口传染,主要传播环节是被含有包囊的粪便污染的水源。居民点的水源被污染常酿成该地区的爆发流行和异乎寻常的高感染率;其次是手指、食物或用具的被污染;卫生习惯不良的带虫者是危险的传染源,蝇及蟑螂等昆虫也能对包囊起一定的传播作用。由于缺乏有效的获得性免疫,患过阿米巴病的人仍是易感者。易感性与性别,年龄无相关性,流行统计中的男性高发现象,多与生活习惯和职业等因素有关。
防治原则
根据溶组织内阿米巴须通过宿主粪便排出的大量包囊污染水源食物而传播的特点,其防治措施应侧重于几个方面:
1.查治病人和带虫者以控制传染源,特别要发现和治疗从事饮食工作的包囊携带者及慢性患者,必要时应予鉴别虫种,决定治疗对策。抗虫治疗目前以甲硝咪唑(灭滴灵metronidazole)为急性阿米巴病(包括不同部位的脓肿)的首选药物,口服吸收良好,副作用少,但到达结肠浓度偏低,单纯用于治疗带虫者的效果并不理想。根治肠阿米巴病应配伍用喹碘方(chiniofon)、碘氯羟喹(vioform)等对肠腔型阿米巴有效的喹啉类药物。氯喹亦为治疗肠外阿米巴病的有效药物。中药鸦胆子仁、大蒜素、白头翁等也有一定疗效,且副作用小。
2.管理粪便,保护水源为切断阿米巴病传播途径的主要环节。因地制宜进行粪便无害化处理,杀灭其中包囊,并严密防止粪便污染水源,应是防制阿米巴病的关键措施。
3.注意饮食饮水卫生,养成良好个人习惯,消灭害虫,搞好环境卫生,防止病从口入,均属保护易感人群的有力措施。
第二节 其它消化道阿米巴
寄生人体消化道的阿米巴除溶组织内阿米巴的复合种群外均为腔道共栖原虫,有些仅偶然寄生人体,一般不侵入组织,但在重度感染或宿主防御功能减弱时亦可产生不同程度的粘膜浅表炎症,或伴随细菌感染而引起腹泻或其它肠功能紊乱。对这些非致病或机会致病的肠道寄生原虫,必须与致病的溶组织内阿米巴鉴别,通常不需治疗,对药物敏感性也常有不同。动物学分类多数归于内阿米巴属(Genus Entamoeba),其它常见的属有嗜碘阿米巴属(Genus Iodoamoeba)和内蜒属(Genus Endolimax)。原属于内阿米巴科的双核内阿米巴属(Genus Dientamoeba)现已作为一种无鞭毛的阿米巴样鞭毛虫而归属于动鞭纲(Class Zoomastigophorea)中的滴虫目(Order Tichomonadida)。以下简述其中的常见种类。
一、哈门氏内阿米巴
哈门内阿米巴(Entamoeba hartmani von Prowazek,1912)形态与溶组织内阿米巴极其相似而体积较小,曾一度被认为是它的共栖小宗。后经研究从形态特征、生长代谢、免疫特性、药物敏感及致病毒力等方面提出证据,确认为独立种。滋养体直径3~12µm,包囊4~10µm。滋养体与包囊的细胞结构和胞核等特征,除大小外酷似非侵袭型的溶组织内阿米巴,糖原泡不明显,拟染色体细小,亦呈棒状小体,也熟包囊也有4个核。本虫不致病,传播及分布与溶组织内的阿米巴相似,常并存感染,仅感染率较低,对硝基咪唑类药物也不甚敏感。病原检查时识别该虫的单纯感染具有缩小防治范围的实际意义。流行病学调查中,测量包囊大小,以10µm为界线,可与痢疾阿米巴包囊相区别(图10-4)。大小在界线交叉范围者鉴别十分困难,有时须借助血清学辅诊。

图10-4哈门氏内阿米巴
二、结肠内阿米巴
结肠内阿米巴(Entamoeba coli Grassi,1879)是人体肠道最常见的共栖原虫,不致病。滋养体直径10~50µm,略大于溶组织内阿米巴。胞质呈颗粒状,内外质不分明,活动迟缓。内质含大量细菌、酵母菌及淀粉粒等食物泡,但不含红细胞。具有鉴别意义的核,经铁素木素染色后可见核周染粒粗细不匀,排列不齐,核仁稍大,经常偏位。包囊球形,直径10~30µm或更大,明显大于溶组织内阿米巴包囊。胞核4~8个,成熟包囊偶有超过8个者。核亦能在未染色的活体中见到。未成熟包囊常有较大的糖原泡。拟染色体常不清晰,似碎片状,两端尖细不整。生活史与流行情况与溶组织内阿米巴相似,成熟包囊经口感染宿主,除人外,鼠、猪、犬等动物肠内亦发现有寄生。呈世界性分布,在我国与溶组织内阿米巴平行分布,感染率高于后者。发现结肠内阿米巴时有必要继续寻找痢疾阿米巴。(图10-5)

图10-5 结肠内阿米巴
三、微小内蜒阿米巴
微小内蜒阿米巴(Endolimax nana Wenyon et O'Connor,1917)为寄生人、猿、猴和猪等结肠腔的小型阿米巴,滋养体平均直径约10µm,外形、大小很像哈门氏阿米巴,但核型特殊。染色后的胞核可见粗大而不规则的核仁,占核直径的1/3~1/2,常偏于一侧。由于缺核周染粒,核膜显得极薄,与核仁之间有清晰的空隙和相连的核丝。包囊椭圆或类圆形,平均大小9µm,色线灰,不易着染而常易辨认。成熟包囊也有4核,缺拟染色体,偶见形状不一的糖原泡。该虫也不致病,重感染或特殊情况下偶有引起急性或慢性腹泻。该虫对甲硝咪唑类药物敏感。(图10-6)
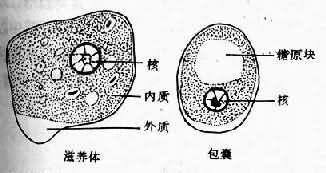
图10-6 微小内蜒阿米巴
四、布氏嗜碘阿米巴
布氏嗜碘阿米巴(Iodamoeba butschlii von Prowazek,1912)以包囊期具有特殊的糖源泡而得属名。虫体稍大于微小内蜒阿米巴,滋养体6~25µm(平均在12µm上下),伪足宽大,不吞噬红细胞,可见1~2个糖原泡。经铁素木素染色后核明显,其特征为中央有粗大的核仁,外围为一层染色较浅的微粒所包绕。核染色质粒纤细,常在核膜与核仁之间形成圈围。包囊呈不规则的卵圆形,直径6~16µm(平均约10µm),但多变异。突出的特点是含有圆形或卵圆形边缘清晰的糖原泡,常把核推向一边。核一般仅一个,核内染色质粒常聚集于核仁一侧呈新月状。碘染标本中,糖原泡呈现棕色团块,而在未染色或铁素木素染标本则为泡状空隙。特殊的糖原泡和核构造是鉴定本虫的主要依据(图10-7)。
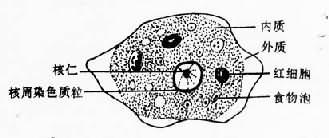
图10-7 布氏嗜碘阿米巴
五、齿龈内阿米巴
齿龈内阿米巴(Entamoeba gingivalis Gros,1849)为人及许多哺乳类如犬、猫等口腔齿龈部的共栖型阿米巴,在不注意口腔卫生的人群中感染率很高,常与齿龈部的化脓性感染并存,偶在支气管粘液中繁殖而出现于痰液中。滋养体直径10~20µm,与溶组织内阿米巴相仿,内外质分明,活动频繁, 食物泡常含细菌、白细胞等,偶有红细胞。核仁居中,以二分裂繁殖,不形成包囊,滋养体主要借飞沫或接触传播。近年报道在子宫置避孕器的妇女阴道及宫颈涂片中查见齿龈内阿米巴。迄今尚未能肯定该虫与牙周病的确切关系,曾有报导113例牙科病人中59%查到齿龈内阿米巴,96例有良好口腔卫生的对照者中亦有32%阳性。(图10-8)
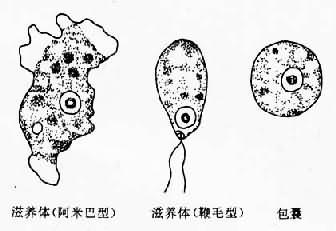
图10-8 齿龈阿米巴
第三节 致病性自生生活阿米巴
自生生活阿米巴种类繁多,广泛分布于水体和土壤内,现已证明双鞭毛阿米巴科中的耐格里属(Naegleria Spp.)和棘阿米巴科中棘阿米巴属(Acanthamoeba spp.)的某些种可侵入人体致病。两者均可引起病程不一的阿米巴脑膜脑炎,迄今报道的病例虽不满200例,却遍及各大洲许多国家,我国也已发现2例,由于全球性分布,可不依赖宿主而生存,病症十分凶险,死亡率极高,已引起广泛注意。
形态与生活史
两类阿米巴均有滋养体和包囊期,胞核都为泡状核形,核仁大,居中。
耐格里属阿米巴,多孳生于淡水中,活动的滋养体呈长阿米巴形大小为7×20µm,常向一端伸出宽大奔放的伪足,另一端较细小为伪尾区,在不良环境中可形成有2根鞭毛的滋养体,此型不分裂也不直接形成包囊;包囊圆形,直径9µm,单核,囊壁光滑有孔,包囊多在外环境形成,组织内不成囊(图10-9),滋养体在35℃下加速增殖,含氯10ppm不能杀死虫体,0.7%盐水可致死。能致病的有福氏耐格里阿米巴(N.fowleri)和澳大利亚耐格里阿米巴(N.australiensis)二种。感染方式主要通过接触污染水体或在游泳池游泳,虫体侵入鼻腔增殖后穿过鼻粘膜和筛状板,经嗅神经上行入脑部寄生。
棘阿米巴多见于污染的土壤和水体中,滋养体为长椭圆形,直径为10~40µm,活动迟缓,体表有多个棘状突起,称棘状伪足(acanthopodia),无鞭毛型;包囊类圆形,双层囊壁,外壁常皱缩,内层光滑呈多边形(图10-10)。现已分离到7个致病种,其中以卡氏棘阿米巴(A.castellanii)为多见。棘阿米巴入侵途径尚不完全清楚,已知可从皮肤伤口、穿透性角膜外伤、损伤的眼结膜或经呼吸道、生殖道等进入人体。多数寄生于脑、眼、皮肤等部位。

10-9 耐格里属阿?, http://www.100md.com(钱宗立)
参见:首页 > 药学版 > 药物手册 > 西药大全一 > 皮肤科、眼科、耳鼻喉科常用药 > 消毒及皮肤科用药 > 碘