23.4 病理生理
由于肥胖的主要发生地点是在机体脂肪组织,所以有关这一节的论述,也就以机体脂肪再重复。
23.4.1 脂肪组织的作用、结构和成分
脂肪组织是结缔组织的一种特殊形式,是机体较大的组织之一。在正常成年男性,它占总体重的15~20%;在正常成年女性,则占总体重的10~25%。它分布于机体某些特定部位,如皮下、网膜、肠系膜、腹膜后、胸腔纵隔障和胸腹浆膜下等处,起着保护和机械支持的作用,并以TG的形式储存能量(这是机体最有效、最紧凑的储能方式),参与糖和脂代谢及体内能量平衡的调节。其中尤以皮下脂肪组织为机体的最大脂肪库,其储脂量约占总体脂量的一半。脂肪组织并不象通常所设想的那样是一种没有多少生活力的组织,它具有与常代谢活跃组织所具有的相同的全套亚细胞结构。在神经内分泌因素影响下,其TG的合成和分解代谢极为活跃。一些研究表明,当机体进行3H以上的有氧代谢活动,其所需能量几乎70%要由脂肪组织来提供,而且随着时间的延长,这一比例还会加大。因此,一定数量的脂肪组织的存在,不仅不是机体的一种额外负担,而恰恰正是机体具有良好身体素质的表现,是机体维持长时间有氧代谢活动的耐力所必需。
脂肪组织主要由脂肪细胞所组成。此外,尚含有少数纤维母细胞和少量细胞间胶原物质。脂肪细胞就处于支持这些细胞及通过 这个细胞群的毛细血管和神经纤维的胶原结构之中。脂肪细胞就处于与毛细血管网一道的较原始的原发细胞;原发细胞分化为储存脂肪的脂肪细胞是受营养和许多内分泌的因素所控制的。
脂肪组织含脂量为60~80%;其所含脂肪都存在于脂肪细胞之内。脂肪细胞的数目,是由遗传因素和幼年时的饮食因素所控制;而中、晚年时的摄能状况,则主要影响已有脂肪细胞体积的大小。当前一般认为,正常人全身脂肪细胞数为26.8±1.8×109;但这只是一项参考数值。人体脂肪细胞较为准确的正常数值之所以迄今尚未确定,这是因为:①用目前的方法,一个脂肪细胞一定要在含有最少量的脂质时才能与其它结缔组织细胞相区别而能以作出鉴定;②目前所用方法在计算机体脂肪细胞总数时,假定全身各部脂肪细胞的平均直径是相同的,这又与客观事实不相符合;③因雌激素水平 的不同,正常男性与女性,不仅其体脂含量而且其脂肪细胞数也是不同的,单一数值显然难以代表。

图23-2 脂肪代谢的某些特征示意图
脂肪细胞是机体最大的细胞,正常人皮下脂肪细胞的平均直径为67~98um,其每一肪细胞的平均含量约0.6ug。含脂质的脂肪细胞在构造上不同于其它细胞的最大特点,主要是由TG组成的一个的中心脂肪滴,将细胞推移到一旁,至少有两个脂肪区域,一个是由许多快速转换的细胞质颗粒所组成,另一个则是前者的最终储存库,即转换缓慢的中心性大脂肪滴。脂肪细胞的主要功能是合成、运转和储TG。这些功能受神经、激素和酶系所控制,并以FFA形式释放入血液,以供其它组织的能量需要。脂肪细胞对TG的储存和释放,完全取决于机体营养与代谢的需要;脂肪代谢的某些特征,如上图所示(图23-2)。葡萄糖是通过胰岛素调节。胰岛素不仅促进清除因子脂酶的活性增加,从而增强血浆中脂肪向脂肪细胞内转移;而且还促进脂肪细胞对葡萄糖的摄取以及对TG的合成。所以,血浆中胰岛素的浓度,显然是决定脂肪组织中脂肪积聚的重要因素。
在机体能量出超的条件下,脂肪组织内所储存的TG经激素敏感的脂酶的作用而释放FFA;从脂肪细胞内弥散出来的FFA与作为载体的白蛋白结合或直接进入其他细胞,以供其他组织的能量需要。现在已知,机体大多数组织均可利用FFA,在许多组织中FFA还是一种优先的能源物质。体内绝大多数的激素,如肾上腺素、去甲肾上腺素、生长激素、促肾上腺皮质激素、促甲状腺激素、黄体生成素、催乳素、胰高血糖素及肠促胰液肽等等,都属于促进脂肪组织释放FFA的激素。
23.4.2 脂肪组织中脂肪储存与释放的机理
在正常情况下,脂肪组织中的脂肪含量极其恒定,这意味着控制脂肪合成和储存的反应与控制脂肪分解和动和的反应处于动态平衡之中。为了弄清肥胖的形成,需要复习一下脂肪组织储存脂肪的调节过程(图23-3)。
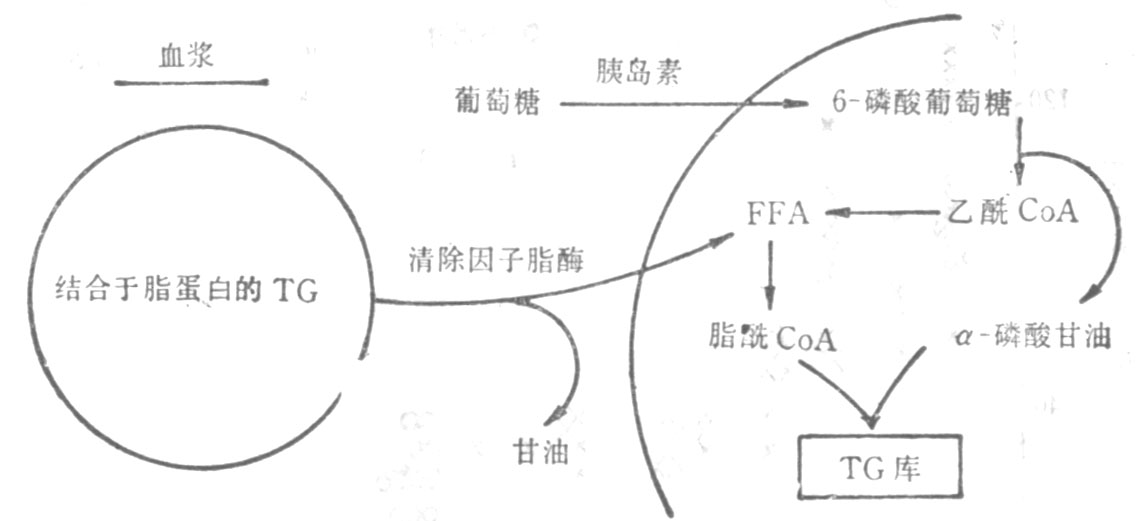
图23-3 脂肪组织中储脂途径示意图
由上图可知,脂肪组织储存脂肪是通过以下两条途径:第一,血浆中的脂肪转移进入脂肪细胞。结合于β-脂蛋白和乳糜微粒的肝源性和肠源性的TG,经秘细血管内皮细胞表面之清除因子脂酶的水解而成为甘油和FFA;接着在酶的调节下FFA进入脂肪细胞,并重新合成TG而储存起来。为了这个途径能顺利和有效地进行,葡萄糖的摄取和代谢是必需的因为分解的激素,故统称为脂解激素;它们通过刺激脂肪细胞表面的特异感受器,激活与膜结合的腺苷酸环化酶,使细胞内环腺苷酸(cAMP)的浓度增加,进而激活素敏感的脂酶,于是导致FFA的释放。第二,胰岛素则为抑制脂肪分解的激素;它以下四种方式限制FFA的释放:①抑制腺苷酸环化酶;②激活cAMP磷酸二酯酶,从而促进cAMP的分解;③减慢FFA从脂肪细胞外流;④促进脂肪细胞对葡萄糖的摄取和对FFA的酯化。其所涉及的具体反应如下图所示(图23-4)。
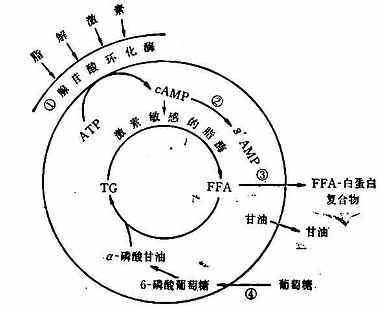
图23-4 脂肪细胞中脂肪的动员
脂肪细胞的数目,年龄愈小愈多;在20岁以后所发生的肥胖,则几乎全由于原有脂肪细胞的肥大所致。这说明,除了遗传因素之外,儿少时期的能量入超,也是一部分人之所以有较多的脂肪细胞的原因。
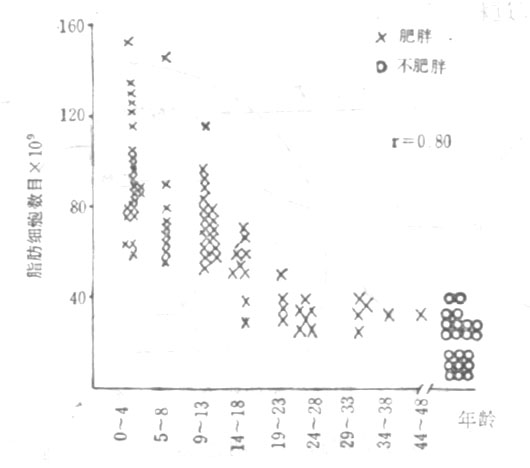
图23-5 肥胖与不肥胖者的脂肪细胞与年龄的关系
由上可见,脂肪组织对脂肪的储存与动用,是在营养、激素和酶系的控制下进行的;而胰岛素在调节这两个过程中所起作用成为重要。
23.4.3 肥胖过程脂肪组织的变化
当肥胖发生和发展时,随着体脂积聚,脂肪细胞含脂量渐增,其体积也明显扩大。以皮下脂肪为例,每一细胞的平均含脂量可由原来的0.6ug增至0.91~1.36ug,其平均直径则可由原来的67~98um增至127~134um。尤其当肥胖的发生与发展过程是缓慢而持续时,有仅可发生更为肥大的脂肪细胞,而且通过脂肪组织的毛细血管内皮细胞表面还可含更多高活性的清除因子脂酶。同时,某些肥胖者还会具有数目较多的脂肪细胞,其全身脂肪细胞数可达77.0±13.5×109。无论是脂肪细胞体积的扩大或脂肪细胞数目的增多,或者是两者兼而有之,其结果都是机体脂肪组织的扩增。
Salans等对78名肥胖者和21名非肥胖者作了对比观察,结果(图23-5)表明:发生肥胖婴儿期明显肥胖、青春发育期肥胖,有这样长期持续肥胖史的成年肥胖者,往往是脂肪细胞的肥大和增生同时并存,其治疗难度也就更大不仅不易取得疗效,而且还能于巩固疗效。据Lioyd报道,有这样长期持续肥胖史的肥胖者,在整个成年肥胖者中占1/3;而其余的2/3,则分别发生于一生中各个有生理性增加体脂的阶段。而Abrham等对10~13岁肥胖和正常儿童各100名(男女对半)作了长期的对比观察,历时20年,结果肥胖儿男性43/50、女性40/50变成肥胖成人,正常儿则男性21/50、女性9/50变成肥胖成人。由上可见,从胎儿时期开始,直至一生中各个有生理性增加体脂的阶段,及早地采取措施预防肥胖,也就至关紧要。
23.4.4 TBAT之状况与机体胖瘦的关系
在人和多数哺乳动物(特别是婴儿和新生仔畜)的颈、肩、腋窝和背部肩胛间,还存在一种特殊类型的脂肪组织。它的结构特点是在脂肪细胞中除含大量分散的TG滴之外,还含有大量线粒体,血液供应丰富,由于其线粒体中细胞色素多,而使之呈现棕色,故通常也就称之为棕色脂肪组织(IBAT)。与其相对应,前述之普通脂肪组织则就称之为白色脂肪组织。
IBAT的主要功能是产热。成年机体遭受刺激,通常可通过战栗和化学两种途径产热;可是,婴儿及新生仔畜由于还不会通过战栗途径产热,其所需之热几乎全要靠化学途径来供应。除Na+-K+泵外,IBAT就是机体化学产热的又一重要装置;在某些小动物甚至其所需总热量的一半是由IBAT所提供。故一定数量质量的IBAT的存在,对机体适应寒冷刺激具有重要意义。IBAT的产热作用,是在交感神经与一种分子量为32000的蛋白质类脱偶合因子的控制下进行的。交感神经有丰富的神经末梢支配这种组织;而这种组织的呣粒体中则有大量的为其所特有的脱偶合因子的存在。寒冷刺激引起交感神经兴奋,其末梢就释放儿茶酚胺,于是就通过cAMP而使激素敏感的脂酶活化,促使TG水解为FFA。这时,又由于该组织中有大量细胞色素及其所特有的脱偶合因子的存在,同时高水平的FFA亦有脱偶合作用,致使FFA被氧化,但偶合则很少,从而能量也就很少产生ATP,而大部分以热能的形式放出。再加上,脱偶合结果导致了ATP~ADP比值的减少,而较多的ADP又使氧化磷酸化及呼吸作用加快,这样也就更使热生成作用增强。
晚近通过对IBAT产热机理的深入研究,发现IBAT之数与质的状况与机体的胖瘦之间有着密切的相关。就目前所知,消瘦、正常、肥胖三者相比,除了机体所含白色脂肪组织明显顺序递增而IBAT明显顺序递减外,其IBAT线粒体中所含的特有的脱偶合因子的数量和活性也明显顺序递减。正常者由于其白色脂肪组织与IBAT之间比例为平衡,而且脱偶合因子的数量和活性也较适中,能对产热和产生ATP进行有效的调节,故在通常情况下能使机体能量得以维持正常的平衡,即能使之不过多以热生成的形式而被消耗,又能使之不过多转化为脂肪而在体内积聚。至于消瘦者,则往往是因为其IBAT含量较多,以及其脱偶膈因子的数量和活性较高,致使其能量较多地以热生成的形式被消耗了,所以在一般情况下也就难以有较多的体脂积聚;反之,肥胖者则由于其所含IBAT量少或功能障碍,致使产热这一有效的调节方式失灵,所以引起能量的入超,其大部分就转化为脂肪而积聚起来。
, 百拇医药