二、近交后引起的变化
(一)近交可以增多纯合性,也就是降低杂合性(Reduction of the terozygosity)
自然界的动植物在遗传上一般都是杂合子体,故最初培育近交系动物都是取源于杂种,换句话说也就是取源于异型接合体动物。动物的遗传纯度取决于交配动物之间遗传关系的远近。因此近交系动物通过长期近亲交配而建立的。通过纯育后,动物的杂合性逐渐降低,因而纯度逐渐增高,使基因位点变为纯合子,使它们的表现型趋向一致性,如A与a基因频率相同,但基因型分配变化,经过一定代数的兄妹交配,则不再有杂合子,而获得遗传稳定的AA与aa近交品系,见表3-1。同胞兄妹或亲子交配连续20代以上就培育成近交系,为了保持该系的遗传特点,必须继续近交,并不得因突变或遗传漂移而丢失。
表3-1 近交引起的基因纯合情况
P(A)=0.5 p(a)=0.5
|
代数 |
各 种 基 因 的 比 例 | ||
|
纯合子(野鼠色)AA |
杂合子(野鼠色)Aa |
纯合子(黑色)aa | |
| 原 始 种 群 |
1/4 |
2/4 |
1/4 |
| 兄妹交配下一代 |
5/16 |
6/16 |
5/16 |
| 兄妹交配下二代 |
11/32 |
10/32 |
11/32 |
| 兄妹交配下三代 |
24/64 |
16/64 |
24/64 |
| 兄妹交配20代以后 |
50/100 |
0 |
50/100 |
实验动物交配繁殖中的一些概念和符号。
1.纯合子:同源染色体的相对位点上具有两个相同的基因称为纯合子(Homozygote),如AA(显性纯合子),aa(隐性纯合子)。
2.杂合子:两个基因不相同时,即由一个显性基因和一隐性基因结合而成的杂合子交(Heterozygote),如Aa。
3.杂合子的a位点:即指a位点上两个位点为A/a。
4.纯合子的等位点:用A/A或a/a表示。
5.显性突变位点用D表示,隐性突变位点用r表示。
6.D和r位点在繁殖制度中被称作重要位点(Loci of interest),因为它们是控制位点分离的。
7.D位点上的等位基因为A/D,r位点上的等位基因为A/r。
8.以在一个位点上的等位基因A/A、a/A、a/a三种基因型进行组合将产生九种交配形式,可归纳为以下四类交配系统。
(1)纯合子交配(Incrosses):为亲代相同的显性纯合子交配,如A/A×A/A和a/a×a/a。
亲代:♂AA×♀AA 子代:基因型AA,表现型A。
| 精子 卵子 |
A |
| A |
AA |
(2)杂交(Crosses): 为亲代不相同的纯合子交配,如A/A×a/a和a/a×A/A。
亲代:♂AA×♀aa 子代:基因型Aa ,表现型A。
| 精子 卵子 |
A |
A |
| a |
Aa |
Aa |
| a |
Aa |
Aa |
(3)回交(Backcrosses):为亲代显性纯合子与杂合子交配。如A/A×a/A;a/A×A/A;a/A×a/a和a/a×a/A
亲代:♂Aa×♀AA 子代:基因型:1AA,1Aa 表现型:A,均为显性野鼠色。
| 精子 卵子 |
A |
a |
| A |
AA |
Aa |
| A |
AA |
Aa |
(4)自交或称互交(Intercrosses):亲代为两个杂合子交配:如a/A×a/A。
亲代:♂Aa×♀Aa 子代:基因型:1/4AA,2/4Aa,1/4aa。表现型:3/4A,1/4a。
| 精子 卵子 |
A |
a |
| A |
AA |
Aa |
| Aa |
Aa |
Aa |
C57BL黑色a,C3H野鼠色,二者皆为纯合子。二者杂交:

种系中纯合性的程度通常以近交系数(F)表示,所谓近交系(Coefficient of inbreeding)也就是一个种系的个体中,两个等位基因具有相同来源的概率,通常以0~1之间的尺度来表示。F=0为完全杂合,F=1表示完全纯合。F值随近交繁殖连续的代数而递增。增加的比率决定于交配动物亲缘关系的密切程度。根据Feiconer(1960)的研究,认为全同兄弟姐妹交配,近交系前几代数值不恒定,如前四代近交系数上升率分别为28%、17%、20%和19%,以后每代上升率就恒定为19.1%。故Felconer提出一个便于计算的公式:Fn=1-(1-△F)n。n表示近交代数,△F是每进一代的近交系数上升率。例如繁殖了10代,△F为19%,代入公式,则
F10=1-(1-0.19)10=1-(0.81)10=1-0.1216=0.8784=87.84%
也就是说繁殖到第10代时,纯度可达到87.84%,还有12.16%是杂合的。由于交配方式不同,其△F也各不相同,同胞兄妹交配△F为19.1%,同父异母交配11.0%,回交(亲子交配)19.1%,堂兄妹交配为8%。全同胞兄妹或亲子交配前20代的近交系数计算状况可见表3-2。
表3-2 Felconer近亲系数表
|
代 数 |
F |
代 数 |
F |
|
1 |
0.250 |
11 |
0.908 |
|
2 |
0.375 |
12 |
0.926 |
|
3 |
0.500 |
13 |
0.940 |
|
4 |
0.594 |
14 |
0.951 |
|
5 |
0.672 |
15 |
0.961 |
|
6 |
0.734 |
16 |
0.968 |
|
7 |
0.785 |
17 |
0.974 |
|
8 |
0.826 |
18 |
0.979 |
|
9 |
0.859 |
19 |
0.983 |
|
10 |
0.886 |
20 |
0.986 |
注:近交系不改变频率。
(二)近交可将群体分离为不同基因型的品系(Splitting of the Population into Lines of Different Gentypes)
原始种群经过近交分为若干近交系,在不同等位基因的不同位点上变为纯合子。原始种群的小鼠兄妹交配连续20代之后分离出两支等位基因A和a的近交品系:
1.等位基因A的品系:只生产野鼠色后代。
2.等位基因a的品系:只生产黑色后代。
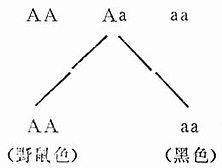
近交品系动物的获得,可使原来杂合子的动物增加纯合性(Homozygosity),从而提高基因型的稳定性,获得想要的品系,实验者可以从动物的特定性状中进行选择。
(三)近交可引起近交衰退(Inbreeding Depression)
亲缘接近的交配所产生的后代常常会出现生长、成活、生育、抗病、适应环境等能力的减退,这种现象称之为近交衰退,如何克服和解决这个问题,是培育近交系动物的关键所在。由于近交衰退的缘故,从一个供繁殖的种群培育和建立一个近交系时,往往需要从杂交群或几个近交系开始才能成功,而且的近交几代后,有些系可以因生育或生活率低而断绝;留下的受近交衰退影响较小的后代有些可以维持下来,而另外的也可能最后被淘汰。也由于同样的原因,过去建立的某些近交系,现在只有某些亚系还仍然存在,有些不能全部存活,甚至难以维持。
近交衰退发生的原因是多方面的,从遗传学的角度解释主要有两点:
1.有害的隐性基因的暴露。一般病态的突变基因绝大多数都是隐性的,所以处于杂合状态时是不表现出病态或不利的性状。这些有害基因的作用可被显性的杂合子等位基因所掩盖,但经过一段近亲繁殖,纯合的基因(纯合子)比例渐渐增多,于是有害的隐性基因相遇成为纯合子而显出作用,出现了不利的性状,对个体的生长发育、生活和生育等产生明显的不利影响。例如杂种动物所带有的不育的隐性基因往往被其显性的等位基因所掩盖,而不表达其不育的性状,但由于纯育,动物的纯合性逐渐增高,不育的现象也就表现出来了。
2.多基因平衡的破坏。个体的发育受多个基因共同作用的影响,虽然其中每个基因的作用效应微小。对环境适应较好的野生或杂交动物,由于自然选择的作用有利于保存那些生物适应能力较强的基因组合具有平衡的多基因系统,近交繁殖往往会破坏这个平衡,造成个体发育的不稳定。
近交衰退往往在近交培养过程中的最初若干世代中表现出来,以后经过一定的人工选择,带有纯合有害基因的动物被逐渐淘汰,或者由于无意识地保留了一小部分的杂合性,经过5~10代左右的培育繁殖,后代中生育与生活力可以逐渐稳定,不再下降。
由于近交系动物是采用近亲繁殖方式,因此容易引起生活力的降低、生长繁殖力退化、抗病力降低等近交衰退变化,为了防止这些问题产生,应注意如下几点:
1.为防止种群传代终断,可采用回交方式(即父×女或母×子)而不计算近交代数,这样可以维持种群不致在繁殖中终断。
2.饲料营养的保证很重要,在一般传代至5~7代时会出现生命力下降,生长繁殖退化,或出现产仔畸形等情况,因此应在饲料中适当增加营养成分以保证子代生长发育正常。
3.在传代中注意检查子代的情况选优去劣,选择体质健康、生殖力旺盛的后代而不能选择体弱繁殖力差的动物。
4.近交到6~7代时,可出现致死的有害基因,而且这种致死基因随近交代数增加而不断升高,可引起个体死亡,为此,培育近交品系,开始不必用同窝兄妹交配,用同父异母,或同母异父的兄妹交配,如待近交品系建立后再改变致死基因则很困难。
, http://www.100md.com