二、抗独特型抗体在医学中的应用
抗独特型抗体不仅对于免疫应答调节的理论研究具有重要意义,而且在传染性疾病的预防、自身免疫性疾病发病机理研究和恶性肿瘤的治疗等应用研究中也具有重要的价值。
(一)抗独特型抗体与疫苗
抗独特型抗体中Ab2β作为抗原的模拟物,可代替病原体的抗原,诱导机体产生抗病原体的特异性免疫应答,即所谓抗独特型疫苗(anti-idiotype vaccine)。目前,抗独特型疫苗研究仍处于动物实验研究阶段,疫苗的范围主要有病毒、细菌和原虫等引起的传染病(表9-2)。
1.抗病毒的抗独特型疫苗 针对病毒感染的抗独特型疫苗是近年来较活跃的研究领域。Reagan将狂犬病毒糖蛋白特异的单克隆抗体免疫动物,制备出抗Id抗体,将这些抗体注射小鼠后,可诱导出具有病毒特异性中和活性的抗体应答。Kennedy等用抗HBsAg抗体制备出抗独特型抗体,经亲和层析纯化后注射于BALB/c小鼠,诱导产生针对HBsAg的体液免疫应答。
Uytdehaag等用对Ⅱ型脊随灰质炎病毒特异的保护性单克隆抗体(Ab1)免疫BALB/c小鼠,制备出单克隆Ab2,用Ab2免疫BALB/c小鼠后,可诱导出Ab3,产生抗脊髓灰质炎病毒的保护性免疫,这种单克隆Ab2可用于预防Ⅱ型脊髓灰质炎病毒的感染。Tanaka等在BALB/c小鼠中用抗新城疫病毒(NDV)血凝素的抗体诱导出抗Id抗体,在无病毒抗原存在的情况下,这种抗Id抗体可刺激小鼠产生抗NDV抗体。Lathey用单纯疱疹病毒(HSV)糖蛋白D特异的McAb制备了抗Id抗体,并在小鼠中诱发对HSV的免疫应答。用人工合成HIV包膜抗耕牛的735-752氨基酸段的多肽免疫黑猩猩制得Ab1,再用Ab1免疫家兔制备Ab2,将Ab2免疫小鼠获得Ab3,实验表明,Ab3能与HIV多肽抗原发生特异性结合。
图9-10 HBsAg抗Id的实验研究(Kennedy,1984)
2.抗细胞菌和原虫的抗独特型疫苗 抗Id抗体也可用于诱导针对细菌和原虫的保护性免疫应答。例如,用Ab2诱导动物针对肺炎球菌、大肠杆菌(K13)、血吸虫和罗德西亚锥虫等病原体的保护性免疫。其Ab2的制备和研究方法与病毒性抗Id疫苗研究类似。
抗Id作为疫苗有以下优点:(1)不含有传染性物质;(2)可大量制备抗Id的McAb;(3)某些保护性抗原决定簇为碳水化合物,不能用DNA重组技术获得,而某些抗Id可模拟碳水化合物抗原决定簇;(4)能诱导比灭活病毒更为有效的T细胞应答;(5)诱导抗原决定簇特异性的抗Id疫苗。应该看到,抗Id疫苗的免疫应答水平一般较低,大剂量鼠源性免疫球蛋白可能刺激机体产生抗异种蛋白的免疫反应以及过敏反应。因此,在临床实际应用中,制备人源性抗Id疫苗,应该看到,抗Id疫苗的免疫应答水平一般较低,大剂量鼠源性免疫球蛋白可能刺激机体产生抗异种蛋白的免疫反应以及过敏反应。因此,在临床实际应用中,制备人源性抗Id疫苗,提高抗Id疫苗的免疫原性,以及细胞工程和基因工程相结合等方面有待进一步深入研究。
(二)抗独特型抗体与自身免疫性疾病
自身免疫病是机体产生对自身成份的免疫应答并造成病理损害而引起的一系列疾病,常与自身隐蔽抗原的释放、自身反应的T细胞克隆激活以及由于具有与自身交叉反应抗原的病原体感染有关。随着免疫网络学说和抗独特型抗体研究的发展,进一步提示了抗独特型抗体与某些自身免疫性疾病的发生有关。
表9-2 抗独特型疫苗的研究
| Reagan等(1983) |
| Sharpe等(1984) |
| Kennedy等(1984,1986) |
| Uytdehaag等(1985) |
| Tanaka等(1986) |
| Lathey等(1986) |
| 米力等(1993) |
| McNamara等(1984) |
| Stein等(1984) |
| Ward等(1987) |
| Naussenzweig等(1980) |
| Sack等(1982) |
| Grzych等(1985) |
1.抗独特型抗体与重症肌无力 1971年,Bartels等合成了称之为Bis-Q(trans-3,3`-bis[(α-trimethyl ammonio methyl)azobenzene]的化合物,发现Bis-Q可模拟乙酰胆硷(acetylcholine,Ach)与乙酰胆硷受体(acetylcholine receptor,AchR)结合。继而,Erlanger将Bis-Q与蛋白载体交联,制备了Bis-Q特异性抗体。1982年Wassermann等用抗Bis-Q抗体免疫家兔,由于抗Bis-Q能模拟AchR,免疫后产生的抗独特型抗体可模拟抗AchR,结果在接受抗Bis-Q免疫的5只家兔中,在第1次加强免疫后就有4只产生了实验性重症肌无力(myasthenia gravis,MG)的症状,但第2次加强免疫后MG症状又有所减轻。他们认为,第1次免疫后产生了抗AchR,阻断了运动终板处Ach与AchR的结合,与MG的症状产生有关。而第2次免疫后,由于注入的抗Bis-Q对血清中抗AchR抗体有中和作用,抑制了抗AchR的作用,使MG症状缓解。重症肌无力患者血清中,除了发现有抗AchR的自身抗体外,还可能存在针对Ach抗体的抗Id抗体,Ach可刺激抗Ach抗体的产生,Ab1可变区的Id又可刺激抗Id抗体产生,Ab2作为Ach的模拟和换封闭了神经肌肉接头的AchR,从而导致神经-肌肉接头功能障碍,发生MG症状。
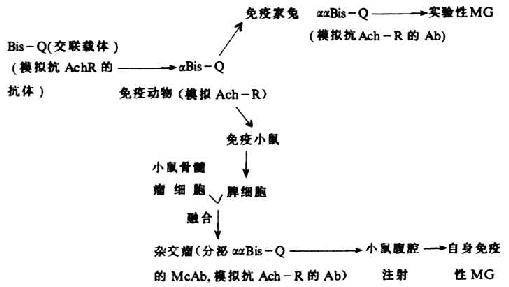
图9-11 Bis-Q、αBisQ和ααBis-Q与实验性MG发生的关系
1984年,Erlanger等又进一步以α-Bis-Q为抗原免疫小鼠,制备了分泌ααBis-QId McAb的杂交瘤细胞株,将此杂交瘤细胞注入到小鼠腹腔,小鼠发生自身免疫性MG,这是由于杂交瘤所分泌的αα-Bis-QId McAb可模拟抗AchR抗体的作用,与Bis-Q的封闭作用相似。
2.抗独特型抗体与促甲状腺受体病 甲状腺机能亢进(简称甲亢)的病因尚不十分清楚。以往认为,病人血清中存在着抗促甲状腺素受体(thyroid stimulating lormone receptor,TSHR)的抗体,由于该抗体与TSHR的结合可模拟TSH的配体作用,因而刺激产生甲状腺素,导致患都产生甲亢的症状,并将抗TSHR称为长效甲状腺刺激素(long acting thyroid stimulator,LATS)。以后研究发现针对抗TSH的独特型抗体中有一部分应能模拟TSH的作用,能与TSHR结合而刺激甲状腺素的分泌。
进一步研究证实:(1)来自甲亢患者血清中TSHαId可促进甲状腺上皮细胞对碘的摄取;(2)αId作为TSHR的配体,检测到TSHR的分子量为197kDa;(3)αId的Fab段可直接与TSH McAb结合并TSH与TSH McAb的结合,TSH也可抑制αId与TSHR的结合。
3.抗独特型抗体与类风湿关节炎 类风湿关节炎患者血清中存在类风湿因子(rheumatoid tacfor,RF),在来自不同个体的类风湿因子之间存在着不同程度的交叉反应,表明这些RF存在着交叉反应独特型(IdX)。Bona等在研究针对人IgM型RF的IdX时,发现一种特殊的抗独特型抗体,称为epidody,这种抗体不仅能与IgM类风湿因子可变区结合,同时也可与IgG的Fc段结合。Chen等合成了与该种IgM RF可变区IdX中氨基酸序列相同的一段多肽作为抗原,免疫家兔制备抗体,结果该抗体可与IgM RF的轻链和IgG结合,提示IgM RF的轻链和IgG上具有交叉反应性抗原决定簇。以上述实验为依据,似乎存在这样的可能:由RF刺激机体产生的epibody可广泛地与人IgG Fc段结合,所形成的免疫复合物沉积于关节滑膜等部位,导致类风湿性关节炎等自身免疫损伤。此外,有人认为许多自身抗体上具有交叉反应Id,这些IdX与外界微生物感染时诱发产生抗体的Id有交叉反应。当相应的微生物感染机体时,刺激产生特异性抗体以及相应的抗Id抗体,后者可通过与自身抗体可变区上IdX的相互作用,诱发自身免疫应答,从而导致免疫损伤。
(三)抗独特型抗体与肿瘤免疫
表达于肿瘤细胞的肿瘤相关抗原刺激小鼠产生相应的抗体(Ab1),用这种Ab1免疫另一小鼠可诱导产生针对Ab1的抗独型抗体,其中Ab2β(模拟肿瘤相关抗原)注入荷肿瘤的动物或病人可诱导出称之为抗-抗-独特型抗体的Ab3,由于这种Ab3与最初肿瘤相关抗原所诱导的Ab1具有相同的抗原特异性,因此又称为Ab1`。

图9-12 模拟肿瘤相关抗原的Ab2β诱导出Ab3(Ab1`)
采用模拟肿瘤相关抗原的Ab2β刺激机体的抗肿瘤免疫应答,已在许多动物实验中得到证实,包括B淋巴细胞肿瘤、T细胞性白血病、乳腺癌、黑素瘤、小鼠肉疚、结肠癌、胰腺癌等(表9-3)。目前,有几种肿瘤相关抗原抗Id已开始临床I期验证,有些病例显示出一定的疗效。
Lery等(1982、1985)应用淋巴细胞杂交瘤技术,制备了针对病人结节性淋巴瘤B细胞表面免疫球蛋白的αId(4D6),获得一定的疗效(图9-13)。
在实验研究中发现,有的肿瘤相关抗原如黑素瘤相关蛋白聚糖(MPG)难于在动物体内诱导出MPG特异性抗体,但是模拟MPG抗原的Ab2β可诱导出肿瘤相关抗原的特异性抗原,提示Ab2β可能通过打破机体对某些肿瘤相关抗原的天然免疫耐受,从而诱导相体产生抗肿瘤相关抗原的免疫应答。
需要注意的是抗Id在一定条件下可抑制机体的免疫应答。Flood等(1980)报道了由抗IdT细胞所诱导的免疫抑制作用。小鼠纤维肉瘤1591和1316都有很强的免疫原性,在正常情况下移植到小鼠体内后100%被排斥。如先用纤维肉瘤1591效应T淋巴母细胞(含TCR Id)免疫正常同类系小鼠,然后分为两组,分别移植1591和1316肉瘤,结果发现,移植1591组约有2/3小鼠丧失排斥肉瘤的能力,而移植1316组仍100%被排斥,说明TCR Id T淋巴母细胞所诱导的免疫抑制作用是特异性的,其机理可能是αIdT细胞杀伤或抑制了肉瘤特异性T效应细胞。
表9-3 肿瘤相关抗原抗Id临床前研究和临床I期验证(举例)
| 肿瘤相关抗原 | Ab1 | Ab2 | 治疗肿瘤 | 应 用 情 况 |
|
图9-13 抗Id(4D6)治B细胞瘤示意图 (金伯泉 许 辉) 参 考 文 献 1.Bhattacharya-Chatterjee M,et al.Idiotype vaccines against human T cell leukemi-a.II. Generation and characterizization of a monoclonal idiotype cascade (Ab1,Ab2 and Ab3).J Immunol,1988;141:1398 2.Bhattacharya-Chatterjee M,et al.Idiotype vaccines against human T cell acute lymphoblastic leukemia(T-ALL).I. Generation and characterization of biologically active monoclonal anti-idiotypes.J Immunol,1987;139:1354 3.Bhattachary-Chatterjee M,et al.Murine monoclonal anti-idiotype antibody as a potential network antigen for human carcino-embryonic antigen.J Immunol,1990;140:2758 4.Bhattacharya-Catterjee MB et al.Idiotypic antibody immunotherapy of cancer.Cancer Immunol Immunother 1994;38:75 5.Chattopadhyay P,et al.Murine monoclonal anti-idiotype antibody breaks unresponsiveness and induces a specific antibody response to human melanoma-associated proteoglycan antigen in cynomolgus.Proc Natl Acal Sci USA,1992;89:2684 6.Davie JM,et al.Structrual correlates of idiotopes.Ann.Ren.Immunol.1986:4:147 7.Grzych JM,et al.An anti-idiotype vaccine against experimental schistosomiasis.Nature,1985;316:74 8.Herlyn D,et al.Anti-idiotype immunization of cancer patients:modulation of the immune response.Proc Natl Acad Sci USA,1987;84:8055 9.Herlyn D,et al.Anti-idiotype antibodies bear the internal image of a human tumor antigen.Science,1986;232:101 10.Jerne NK,et al.Recurrent idiotypes and internal image.EMBO J 1982;1:243 11.Jerme NK.Towards a network theory of the immune systim .Ann Immunol(Paris),1974;125C:373 12.Kennedy RC,et al.Anti-idiotypic antibody vaccine for type B viral hepatitis in chimpanzees.Science,1986;232:220 13.McNamara MK,et al.Monoclonal idiotype vaccine against Strcptococcus pneumoniae infection.Scicnce,1984;226:1325 14.Mittelman A,et al.Human high molecular weight melanoma-associated antigen(HMW-MAA)mimicry by mouse anti-idiotypic monoclonal antibody MK2-23:induc-tion of humoral anti-HMW-MAA immunity and prolongation of survival in patients with stage IV melanoma.Proc Natl Acad Sci USA,1992;89:466 15.Nisonoff A and Lamoyi E.Impllications of the presence of an internal image of the antigen anti-idiotypic antibodies:possible application to vaccine production.Clin Immunol Immunopathol,1981;21:397 16.Rodkey LS.Autoregulation of immune response via idiotype network interactions .Microbiol Rev 1980;44:631 17.Roitt IM.Encyclopedia of Immunology.lst ed .San Diego:Academic Press ine .1992:723 18.Roitt IM.Essential Immunology.6th ed.Oxford:Blackwell Scientific publications.1988;P35-37,P121-125 19.Roitt IM.et al.Idiotypic networks and their possiblse exploitation for manipulation of the immune response.Lancet,1981;I:1041 20.Sacks DL.et al.Immunization of mice against African trypanosomiasis using antiidiotypic antibobies.J Exp Med,1982;155:1108 21.Sharpe AH,et al.Syngeneic monoclonal anti-idiotype can induce cellular immunity to reovirus.J Exp Med,1984;160:1195 22.Stein KE and Sderstrom J.Neonatal administration of idiotype or anti-idiotype primes for protection against Escherichia coli K13 infection in mice.J Exp Med,1984;160:1001 23.Uytdehaag IGCM and Osterhaus ADME.Induction of meutralizing antibody in mice against poliovirus type II with monoclonal anti-idiotype antibody.J Immunol,1985;134:1225 24.Victor-Kobrin,et al.Structural correlates of a regulatory idiotope.1989;Immunol Rev 110:151 25.Wasserman NH,et al.Anti-idiotypic route to anti-acetyecholine receptor antibodies and experimental myasthinia gravis.Proc.Natl.Acad Sci USA,1982,79(15):4810 26.陈敬贤、马宝骊.抗体的独特型与免疫网络研究进展.国外医学免疫学分册,1987;5:238 校对:2000/08/01 刘娜 , 百拇医药 |