第二节 染色方法
一、本科标抗体法
酶标抗体技术是通过共价键将酶连结在抗体上,制成酶标抗体,再借酶对底物的特异催化作用,生成有色的不溶性产物或具有一定电子密度的颗粒,于光镜或电镜下进行细胞表面及细胞内各种抗原成分的定位。
(一)酶的种类及特点
从理论上讲,用细胞化学方法能显示的酶,均可用于标记抗体,进行ICC染色,但实际上在ICC中所能用的酶并不多。现将常用的几种酶列于表4-1,供选用时参考。
表4-1 免疫细胞化学常用的酶
图4-1 过碘酸盐氧化原理 过碘酸盐氧化法,酶的RZ值≥3时较佳;RZ<3时,糖含量较少 ,游离氨基较多,氧化时酶易发生本身聚合,影响酶标抗体的产量,据报告,适当地控制过碘酸盐溶液及反应条件,几乎所加入的HRP和抗体均形成酶标抗体,其标记率为70%左右。具体步骤为: (1)4mg HRP(RZ=3.0)溶于1.0ml双蒸水中。 (2)加0.2ml新鲜配制的0.1mol/LNaIO4,轻轻摇动混合20min,肉眼可见液体内棕黄变成深绿色。 (3)对0.1mol/L醋酸盐缓冲液(pH4.4)透析,20h,4℃。 (4)调整HRP液体的pH至9.5(一般加入20μl0.2mol/L碳酸盐缓冲液pH9.5),立即加入抗体(IgG)8.0mg/Fab 3.0mg (0.01mol/L碳酸盐缓冲液溶解),轻轻混匀后,置室温2h。PH≤8.5时,抗体的NH2基被氧化生成NH3+,后者不能与CHO基反应,所以,保持pH9.0~9.5非常重要。 (5)加入0.1ml新鲜配制的0.4%NaBH4 溶液,置1~4℃ 2h,以稳定酶标抗体复合物。 (6)经透析等去除未反应的NaBH4 ,避免还原过度 。然后经盐析或柱层析等方法分离酶标抗体(方法同戊二醛法)。 如此制得的酶标抗体,加入终浓度1%牛血清白蛋白(Bovine Serum Albumin, BSA)分装后于-80℃可保存数年;亦可加入6%等量甘油,混匀置-20℃/4℃保存1年左右。上述两种标记法是ICC研究中最常用的酶标抗体制备方法。 3.Maleinmide法 上述酶标抗体(IgG)的分子量较大,对抗体的穿透性有一定影响,而在制备过程中,需经两次纯化,即多聚体与单体及单体与非标记抗体的分离。已知单体(1个HRP分子标记1个IgG分子)的分子量为190~200kD,非标记抗体为146~150kD,用凝胶过滤法很难将二者分开。所以,Sternberger等进行了抗体片段Fab的标记。用植物性蛋白酶---木瓜酶(Papain)水解抗体蛋白,可获得一个无抗体活性稳定的Fc段结晶和两个相同的抗原结合片段(Fab),此Fab段为单价,分子量50kD,HRP标记后,单体分子量为90kD,较容易将单体和非标记的Fab段分离。在此基础上,Imagawa(1982)又进行了改良,引入了N-羟基丁二酰酯(Maleinmide),标记抗体的特定部位--- Maleinmide法。该方法的主要原理是:(1)借助双功能试剂Maleinmide活化HRP,使其具有与—HS 基反应的能力;(2)利用胃蛋白酶水解IgG,IgG重链在近羧基端被切断,这样在绞链区(Hinge region)至少可以保留一个S-S键,从而得到一个具有双价抗体活性的F(ab')2段。该S-S键与抗体活性无关。可以通过加入硫基乙醇(2(β)-Mercapto ethanol, HSCH2CH2OH)使其断裂,被还原成—HS基 。如此双价抗体活性的F(ab')2片段即变为带有—HS 基的单价Fab’片段,后者再与活化的HRP结合,便制成了酶标抗体Fab'。由于空间遮蔽的关系,绞链区即使存在两个—HS基,也只能有一个能与酶结合,另一个—HS基不能与酶反应,即酶与抗体Fab’段结合比例为1:1,因此酶标抗体Fab’段绝大多数是单体,很少形成多聚体。在标记过程中,应用过量的活化HRP,还能避免非标记抗体Fab'段的存在。Fab'段的分子量与Fab段相似(50kD左右),所以酶标后Fab'亦较容易与未标记的Fab'分离。此标记方法多用于酶免疫分析研究,其步骤为: ①6mg HRP溶于1.0ml PBS(pH7.0)中。 ②4mg Maleinmide 溶于0.5ml N, N二甲基酰胺(N,N-dimethylformamide)中。 ③将上述两种液体充分混合,持续搅拌1h,30℃。 ④离心取上清,经0.1mol/L PB(pH6.0)平衡的Sephadex G-25柱层析,收集含有蛋白部分的洗脱液,浓缩,制得活化HRP。 ⑤每1.8mg活化HRP,加入已被还原的Fab'2mg,4℃ 20h持续搅拌。 ⑥ Sephadex G-200/Sephacryl S-200分离酶标抗体Fab'片段,保存同前。 (三)酶标抗体的纯化 上述各种方法制备酶标抗体时,除产生酶标抗体外 ,还有未标记的抗体蛋白、游离酶、酶二聚体及偶联剂等。这些均可使酶标抗体的敏感性下降,故酶标抗体须经纯化后应用。现简介几种常用的纯化方法。 1.多聚体的分离 实验中,不同的试剂形成的多聚体的数量各异,即使同样的实验药物和程序,在不同的时间制备的酶标抗体中,多聚体的数量亦不相同。多聚体较单体含酶量多,与组织的非特异吸附增强,使方法的敏感性下降,故用前必须除去多聚体。以凝胶过滤法分离多聚体的效果较好。 2.非标记抗体的分离 非标记抗体与标记抗体具有相同的免疫学特征,能竞争与组织特异抗原结合,而且亲和力较标记抗体强,优先与组织抗原结合。一般认为每个抗体连结1~2个酶分子,并不影响抗体的两个(Fab)抗原结合点 ,但因存在空间遮蔽现象,一个Fab段与组织特异性抗原结合。非标记抗体则不存在这种现象,两个Fab段均与抗原结合,因而亲合力较强。所以 酶标抗体在应用前须用琼脂糖亲合层析等方法去除非标记抗体。 3.亲合层析 利用蛋白A(Protein A)/琼脂糖-刀豆素A/琼脂糖亲合层析柱能制得较纯的酶标抗体。因为蛋白A与抗体(标记和未标记)间有较强的亲合力,不与HRP结合。而刀豆素A与HRP(标记和游离)间的亲合力非常强,不与抗体结合。据此二者并用,能获得较纯的HRP酶标抗体 。该方法省时,分离能力强(约为凝胶层析的600倍)。但有一定的局限性,因为蛋白A并非与所有种属的免疫球蛋白亲合力均较强,例:不能与羊的免疫球蛋白结合,所以难以用于纯化羊的酶标抗体。而刀豆素A与HRP的亲合力极强,甚至呈不可逆性结合,可使部分酶标抗体的活性丧失。 (四)染色原理及步骤 1.基本原理 酶标抗体与荧光色素标记抗体的染色相同,亦分直接法和间接法。直接法是将酶直接标记在每一抗体上,间接法是将酶标记在第二抗体上,检测组织细胞内的特定抗原物质。间接法所用的第一抗体是对组织细胞内某种抗原的特异性抗体(80%~90%的抗血清系由家兔制得,而绝大数单克隆抗体系由小鼠制备);第二抗体则为第一抗体(家兔/小鼠的IgG)的抗体。所以,只要不同的第一抗体均来自同一种属,同一标记的第二抗体就能用来显示其不同特异性抗原的存在,这样可避免了直接法中标记每一种第一抗体的麻烦,并且提高了方法的敏感度。目前的ICC染色中,以间接法为常用,在此着重介绍间接法的染色程序。 2.染色程序 (1)切片准备:见第一章 。 ①石蜡切片经二甲苯或Hemo-De脱蜡,下行酒精至水。②固定/无固定新鲜组织冰冻切片,室温干燥2h以上。 (2)未固定的新鲜组织切片,丙酮固定20~30min(fretrieval ),简而言之,即用蛋白酶处理,去除固定剂交联造成的空间遮蔽(室温),风干15~30min.;已固定的组织冰冻切片及石蜡切片,根据需要可进行组织抗原的激活。 (3)用蜡笔沿切片周围勾划一道屏障,以避免孵育液流失及孵育过程中切片干燥,同时也能节 省抗体用量,保持切片与抗体的充分接触,风干。石蜡切片(滴少量PBS,以防其干燥)直接进行步骤(6)。 (4)切片经PBS或其它缓冲液漂洗3次,每次2min,溶解除去冰冻切片上的OCT包埋剂。但应用ALP标记抗体时,禁用二甲胂酸钠缓冲液漂洗,因后者可使ALP失活。 (5)根据需要,用甲醇+0.3% H2O2处理切片15~30min(室温),封闭内源性过氧化酶的活性。应用ALP标记抗体时,此步可省略。 (6)PBS漂 洗2min(共两次),移至0.05% Tween-20/PBS(或0.22~1% Triton X-100)中5min(室温)。Tween-20为一种表面活性剂,除具有清洁作用外,还可增加组织的通透性,有利于组织细胞内抗原的显示。 (7)4% Block Ace或0.1%~1%BSA湿盒内孵育15~25min(室温),然后;轻轻弃去孵育液(不冲洗);以阻断组织与抗体的非特异性结合,降低背景染色。 (8)根据需要,可用1/5~1/30正常来活兔/羊血清孵育15~20min(室温,以酶标抗体相同种属正常血清为宜,最好是同一动物免疫前的血清)。或者省略此步,而在稀释酶标抗体时, 加入1%~2%的同一种属正常血清。 (9)轻轻弃去孵育液, 滴加含0.2% BSA/0.05 NaN3/PBS稀释的第一抗体(抗体用量:每张切片一般为50~80μl),湿盒内孵育1~2h(20~25℃);免疫电镜用标本,4℃过夜。 (10)PBS充分冲洗(3次×2min),以去除切片上非特异吸附的抗体。应用不同的第一抗体或同时进行对照标本染色时,需分别冲洗,以防相互污染。 (11)经0.05%Tween-20/PBS 2min后,滴加含0.2%BSA/1%正常血清/PBS稀释的HRP酶标记的第二抗体,湿盒内孵育45~60 min(20~25℃)。 (12)PBS漂洗(3次×2min),此处不需分别漂洗,因所有切片均系同一酶标抗体孵育。 (13)未固定或固定较弱的冰冻切片,此处可轻微固定。首先将切片从PBS移至TBS中3~5min,去除PBS,以防其中的磷酸与后面使用的固定液中的CaCl2形成磷酸钙而沉淀后,然后用Baker氏固定液固定5min(室温)此时轻微固定,既不影响抗原抗体结合,又较有利于保存第一抗体孵育前的组织抗原,尤其适用于单克隆抗体的ICC染色。 (14)呈色:HRP标记抗体的呈色液为0.01%~0.1% H2O2 0.01%~0.05% DAB 0.05~0.1mol/L Tris –HCl(pH7.4)。切片经PBS或Tris-HCl液漂洗后,置上述呈色液内10~15min(室温、暗处),亦可镜下控制显色速度。终产物为棕褐色沉淀。用于电镜观察的标本,呈色3~5min即可,防止DAB终产物向周围扩散,影响超微结构定位。另外,显色液应于用前新鲜配制,避免DAB本身氧化变质。新配制的DAB为无色透明液体。若DAB液已氧化变为紫红色,应换新药重新配制。 (15)将切片置流水中(仅光镜观察)或PBS(电镜标本)内,终止呈色反应。 (16)未固定的冰冻切片,可用1%戊二醛-PBS液加强固定5~10min(室温),流水冲洗。 (17)细胞核轻度染色,以甲基绿和苏木精为常用。前者细胞核呈绿色,与HRP/ALP等终产物对比度尤佳,但不适于微波照射的标本。苏木精是组织学、病理学研究中普遍应用的核染色方法,与HRP、ALP的终产物对比度比较好。 (18)DAB呈色的标本,可以系列酒精脱水、Hemo- De透明、DPX封固。其它物质呈色的标本,在有机溶剂中沉淀物易溶解,褪色,所以用水溶性封固剂如明胶甘油等封固,次日,于盖玻片周围涂少许女士用指甲油,可使切片保存时间更长。 (19)镜检、观察记录同一般形态学研究。 (20)免疫电镜用标本,经OSO4后固定,按电镜标本要求处理(参见本书第七章 )。亦可OSO4固定,喷碳(Carbon Coating),利用扫描电镜观察抗原的存在部位。 3.酶标抗体及发色剂的选择
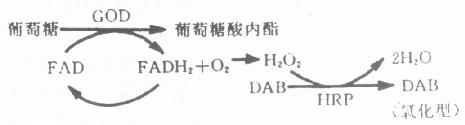
HRP特异底物为H2O2,在分解H2O2过程中,与H2O2形成复合物,无电子供体存在时,反应不再进行,当电子供体存在时,迅速生成水,酶被还原,电子供体被氧化环化,形成苯乙胼聚合体 (图4-2)。在酶反应部位,形成不溶性棕褐色沉淀,与组织对比清晰。
图4-2 DAB反应产物形成过程 HRP催化的酶促反应第一步是特异性的—酶催化底物H2O2,其余反应是非特异的,可用各种电子供体介导,所以选用不同的电子供体,可使终产物呈不同颜色,例如:CN(4—Chloro—1 – N aphthol)为蓝黑色,TMB(Tetramethyl—Benzidine )为深蓝色,AEC(3—amino—9 – ethyl--carbazole)为红色。市售试剂盒所配显色液以AEC居多,室温下较稳定,但不能进行脱水等处理,且时间长易褪色。DAB是广泛应用的电子供体之一 ,较敏感,切片可脱水透明、半永久保存,且终产物具有嗜饿性,经O2O4处理,电子密度增加,适于电镜下确定抗原的存在部位。但是DAB被认为可能具有致癌性,所以应尽量减少吸入和接触次数,最好将DAB制成10倍贮存液,分装于-20℃保存,应用时稀释、过滤。与DAB比较,CN敏感性略差,但因其终产物较局限,很少弥散,光镜观察较为适合。 (2)ALP,是以As-Mx为底物,FB/FR为发色团,生成蓝色/红色不溶性沉淀。ALP标记抗体主要用于内源性过氧化酶含量较高的血细胞、淋巴细胞等的ICC染色。显色液内加入终浓度为2~4mmol/L 的levamisole, 大多数内源性ALP活性可被抑制。通常左旋咪唑(levamisole)以每毫升60~120mmol/L浓度的贮存液-20℃冰箱保存。应用时稀释30倍。为避免反复称取,试剂吸水,As-Mx、FR、FB试剂均应小量分装后-20℃冰箱保存,左旋咪唑能抑制内源性碱性磷酸酶活性。例如:以每次所用显色液30ml计算,分装As-Mx 5~7mg/支、FR 8mg./支、FB7mg/支,每次使用一支,用前稀释30倍,过滤显色 。FR、FB等在光照射条件下易引起沉淀,故显色反应在暗处进行。 (3)GOD,是以葡萄糖为底物的酶,其显色方法为β-D-葡萄糖67mg、NBT6.7mg、PMS(Phenazine Methylsulfate)0.167mg, 0.05mol/L PB(pH8.3)10.0ml,37℃孵育1h。生成蓝色不溶性沉淀。该终产物不溶于有机溶剂,切片可脱水透明,长期保存。但GOD的敏感度较HRP和ALP为低,电子供体少,应用较局限,主要用于两种酶的放大技术,能提高方法和敏感性和特异性。即用GOD和HRP分别标记第二、第三抗体(例第一抗体为小鼠单克隆抗体、第二抗体为GOD标记的兔抗鼠IgG,第三抗体则为HRP标记的羊抗兔IgG),ICC染色,以葡萄糖-DAB作显色剂。基本原理如(图4-3)。在这里HRP是作为第二酶系统,利用葡萄糖氧化时生成的H2O2作底物,催化酶促反应,不受内源性过氧化酶的影响。而且GOD和HRP所标记的抗体是结合在同一抗原位置,所以能良好地显示组织抗原的存在。其主要染色步骤为: ①切片经第一抗的孵育;②漂洗,GOD标记的第二抗体孵育40~60min(室温);③漂洗,HRP标记的第三抗体孵育30~45min(室温);④漂洗,显色、脱水透明观察同前。
图4-3 两步酶催化反应原理 二、非标记抗体酶法 由于酶标抗体存在一些缺点,例如①酶与抗体间的共价连结可损害部分抗体和酶的活性;②抗血清中的非特异性抗体被酶标记后,与组织成分结合,可致背景染色等。为此Sternberger等在酶标法的基础上,发展了非标记抗体酶法。包括酶桥法(Enzyme Bridge Method)和过氧化物酶抗过氧化物酶法(Peroxidase Antiperoxidase Method, PAP法)。现分述如下。 (一)酶桥法 1.基本原理 首先用酶免疫动物,制备效价高、特异性强的抗酶抗体,然后使用第二抗体作桥,将抗酶抗体连结在与组织抗原结合的第一抗体上,再将酶结合在抗酶抗体,经呈色显示抗原的分布。在此过程中,任何抗体均未被酶标记,酶是通过免疫学原理与抗酶抗体结合,避免了共价连结对抗体和酶活性的损害,提高方法的敏感性,且能节 省第一抗体的用量。 2.染色步骤 主要程序如图4-4
图4-4 酶桥法主要染色步骤 (1)切片准备及第一抗体孵育前处理同间接法,第一抗体(假设来自种属A)孵育24h(4℃)。第一抗体的稀释度可高些,使抗体的两个Fab段均与组织抗原结合,较牢固,漂洗时不易丢失。 (2)切片与第二抗体(也称桥抗体,抗种属A IgG抗体)孵育1~1.5h(室温)。应用过量的桥抗体能保证一个Fab段与第一抗体结合,另一个Fab段游离。 (3)切片与抗酶抗体(来自种属A)孵育1~1.5h(室温)。因抗酶抗体和第一抗体均系种属A IgG,具有相同的抗原性 ,所以 桥抗体游离的Fab能与抗酶抗体结合,起到桥的作用,将抗酶抗体连结在与 组织抗原结合的第一抗体上。 (4)切片与酶(HRP 70~100μg/ml,PBS溶解)孵育0.5h(室温),酶与抗酶抗体结合。 (5)显色等与酶标抗体法相同。 酶桥法克服了酶标抗体法的缺点,较好地保护了抗体和酶活性。但仍有其不足:①在酶抗体血清中,含有低亲合力和高亲合力两类抗体,它们作为抗原与桥抗体结合,主要依赖于桥抗体对它的亲合力,而与其本身对酶的亲合力无关,故二者均可被连结在桥抗体上。低亲合力的抗酶抗体与酶结合较弱,漂洗时易解离,使大部分酶(70%左右)丢失,降低了方法的敏感性。②第三步所用的抗酶抗体血清中,亦含有非特异性抗体,其抗原性与抗酶抗体相同,所以能与桥抗体结合,但不能与酶结合,影响组织抗原的显示。为此70年代初,Sternberger(1970)又建立了PAP法,并加以改良,现为ICC研究中常用的方法之一 。 (二)PAP法 1.PAP的制备及特征 PAP复合物是离体制备的HRP抗HRP复合物,它的制备方法较多,现简介其中之一。 (1)制备抗HRP血清:健康雄性家兔(2.0kg以上),可先于足跖皮下注射灭活的卡介苗(共10mg),2周后重复1次,刺激机体免疫系统功能,1周后于背部脊柱两旁皮内多点注射1.0ml乳剂[(福氏完全佐剂,含HRP3.3mg(RZ=3.0) ];间隔2周第2次注射,背部注入含HRP3.3mg的不完全福氏佐剂1.0mg;再间隔2周,第3次注射,2mgHRP(溶于2ml生理盐水中),分别于前后小腿皮下和背部肌肉内多点注射,1周后采静脉血,检查效价。再隔1周第4次加强注射(条件同第3次),一周后检查抗体效价 ,琼脂糖扩散法(HRP0.1mg/ml)为1:128以上时,颈动脉取血,制备血清低温保存备用。 (2)制备PAP复合物:离体条件下,使兔抗HRP抗体与HRP形成可溶性复合物。在制备抗HRP血清时,HRP的纯度要高(RZ≥3),而制备PAP复合物时,HRP的质量要求不高,甚至可用酶的粗制品。 【步骤】 ①取10.50ml抗HRP血清,加7.60ml含7.6mgHRP(1.0mg/ml)的水溶液。 ②混匀,室温1h后,离心16000g 4℃ 15min。 ③0.9% NaCl(冷盐水)溶解沉淀,离心16000g 4℃ 15min,重复4次。 ④加15.3ml HRP水溶液(含HRP30.64mg),室温,持续搅拌。 ⑤用1N HCl及0.1 N HCl调pH至2.3,沉淀物全部溶解变清,立即用1N 及0.1N NaOH调pH至7.2。 ⑥慢慢加等体积的饱和硫酸铵,置0℃ 45min。 ⑦离心35000g 0℃ 15min,沉淀用50%硫酸铵漂洗两次。 ⑧用10.50ml水溶解沉淀,对透析液(48.6g NaCl, 1.5N NaOOCCH3 30ml, 3N(NH4)2SO430ml, 水5.94L)透析,4℃离心,收集上清,加透析液使体积至10.50ml,分装低温保存。 【质量鉴定】 制备的PAP复合物应检测酶和抗体的含量,以鉴定PAP复合物的质量。常用分光光度计检测PAP的复合物在280nm( IgG的最大吸收波长)和400nm(HRP的最大吸收波长)的光密度值(OD值),按下式计算IgG和HRP的含量: 一般HRP/抗HRP的克分子比达1.9时,可分装冷藏于-85℃冰箱,据报告如此冷藏的PAP复合物可保留14年之久,活性无改变。但稀释后的PAP复合物不稳定,4℃可保存2~3周。最好临用前配制。 【PAP复合物的特征】 PAP复合物的形成不同于其它抗原抗体反应,在抗原稍过量时,所有的抗HRP抗体均参与形成可溶性PAP复合物,仅残留少许游离的HRP,而大多数抗原抗体反应需要抗原绝对过量才能形成可溶性复合物。PAP复合物的形状相当稳定,不受抗原量的影响,无论最初加入的抗原抗体过量与否,最终形成的PAP复合物其HRP/抗HRP之比绝大部分为3:2。应用离心沉降、液相扩散等方法分析表明:PAP复合物沉降系数为11.5s,分子量为400~430kD,由此可推算出酶与抗体之比为3:2,即每个PAP生命物由3个HRP分子和2个抗HRP抗体组成,呈五角形结构,3个角为HRP,另两个角为抗HRP抗体。采用H2O2-DAB染色,电镜下已经观察到PAP复合物五角形环状结构,直径平均21nm 。这种结构异常稳定。据报告PAP复合物的抗HRP抗体与HRP结合常数为108,在此可溶性复合物中,即使存在少量游离HRP,亦不影响其稳定性。 2.PAP染色原理及步骤 (1)原理:与酶桥法相似,都是借助桥抗体将酶连结在与组织抗原结合的第一抗体上,所不同的是PAP法将酶桥法的步骤3、4合并为1,用PAP复合物代替,即第三步用PAP复合物孵育切片,故称PAP法。PAP复合物中的抗HRP抗体和第一抗体为相同种属动物的IgG,所以桥抗体能够作为“桥”将PAP复合物连结在第一抗体上。 (2)染色步骤:主要步骤与酶桥法相似,如图4-5。 ①切片准备及第一抗体孵育前处理同间接法;切片与特异性第一抗体孵育同酶桥法。 ②用过量的桥抗体孵育,作用同酶桥法。
图4-5 PAP法主要染色步骤 ③用离体制得的PAP复合物(1:30~300稀释)孵育1~1.5h(室温),使其被桥抗体连结在第一抗体上。亦可用ALP抗ALP复合物代替。 ④显色观察等同间接法。 3.PAP法的评价 PAP法应用比较广泛,特别是近几年,PAP、ABC等试剂盒商品化,在科研和临床病理诊断等中有着广阔的前景。其主要特征和注意事项如下: (1)抗体活性高:非标记抗体酶法最大限度地保存了抗体活性,因为在所有的反应过程中,任何抗体均未被酶连结,避免了标记过程(共价键连结)对抗体活性的损害。 (2)灵敏度高:灵敏度是指ICC方法所能发现最少数量抗原而言。从理论上讲,PAP法应该较间接法敏感2倍以上,因为酶标抗体法中,酶与抗体为1:1标记,而PAP复合物含有3个HRP分子。假设一个第一抗体同一个酶标抗体/PAP复合物结合,则被连结于抗原部位的酶分子数量不同,所以PAP法应较敏感。我们知道组织切片抗原的含量是未知的,所以不能直接在切片上检测方法的敏感度;但可以假设相邻切片抗原浓度相对恒定,采用相邻切片染色,以产生特异性染色所用的第一抗体最低浓度作为方法的敏感度,用信噪比(Signal/moise, S/N)表示。据此,Sternberger(1979)认为PAP法较间接法敏感20~25倍,可进一步稀释第一抗体,以节 省其用量。但实际上PAP法与间接法之间的差异并非如此明显。笔者在实验中注意到,PAP显示阳性的抗原,相同的稀释倍数的第一抗体在间接法中亦呈阳性反应,而时间阴性时,PAP法亦为阴性。其原因尚不清楚,可能与桥抗体和第一抗体结合时,“剩余”(游离)的Fab段少,PAP复合物被连结的相应减少有关 。另外 ,PAP复合物分子量较大(400 KD),冰冻切片时,对组织穿透性远不如酶标抗体(分子量90~180kDa),所以不适于免疫电镜的标本制作。 (3)背景染色低:在酶标抗体法中,被标记的非特异性抗体可与组织成分结合,造成背景染色(图4-6)。从理论上讲,在非标记抗体法,连结抗体中,即使存着非特异性抗体,因其不是抗第一抗体种属IgG的特异性抗体,故亦不能与抗HRP抗体结合,不能把PAP复合物连结在此非特异性抗体 上(图4-7)。当然,PAP复合物内也可能存在些非抗HRP的抗体,假如这部分抗体能够与桥抗体及组织成分结合,但因其不是抗HRP的抗体,故不能与HRP结合,无酶活性。因此说桥抗体的引入,使ICC的特异性得到了双重放大,S/N增大。但也有人认为,过量的桥抗体及PAP复合物等均易与Fc受体结合,致使背景染色增强。实验表明:合适的切片准备、恰当地应用封闭性阻断剂、正常血清以阻断非特异性结合,加之适当地 选择抗体稀释度和抑制内源性酶活性,PAP法和间接法二者的背景均相当低。
图4-6 间接酶标抗体法 背景染色增高原因
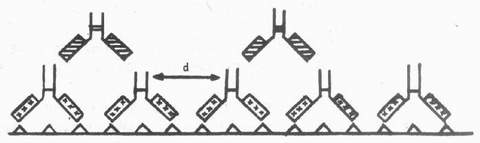
图4-7 PAP法降低背景的原理 (4)假阴性及其处理:应用PAP法显示抗原含理较多的组织或冰冻切片组织抗原保持良好的标本时,可导致阴性结果,这种现象称为假阴性。Vandesand发现:应用PAP法显示大鼠视上核和室旁核内加压素神经细胞分布时,第一抗体(1:200)染色,加压素含有细胞呈强性。如果取材前24~48h,脑室内注射秋水仙素,阻断轴突运输以增加视上核和视旁核加压素含量,同时组织标本经脱水、透明、再水化等处理以增加抗体的通透性,同样稀释度染色,结果却阴性。进一步实验发现:增加抗体的稀释度,开始出现阳性染色,稀释至1:1500时,染色最强,继续增加抗体稀释度,染色强度则下降。Bigbee(1977)认为假阴性是第一抗体用量过高与组织抗原结合过多,使相邻两个伉体的Fc段间的距离(d)恰好是连结抗体的两个Fab段与之结合的长度,于是连结抗体不存在游离Fab段,仅剩无活性的Fc段,所以不能将PAP复合物连结在与组织抗原结合的第一抗体上,结果阴性(图4-8)。因此,借助PAP法研究组织抗原分布,特别是新鲜组织冰冻切片组织抗原保存较好时,第一抗体尽量用高稀释度,避免假阴性。石蜡切片很少发生这种现象,所以PAP法在石蜡切片的ICC观察中更为常用。
图4-8 示假阴性:组织切片上结合过多的第一抗体,两抗体间的距离d 恰好适于桥抗体的两个Fab段与之结合,无PAP复合物结合的位置 (5)桥抗体:也称连结抗体,它的两个Fab段具有相同的结构、性质及与抗原结合的能力。因此,当第一抗体和PAP复合物中抗HRP抗体来自同一种属动物时,桥抗体能同时与上述两种抗体结合,起到桥的作用。为了确保桥抗体的两个Fab段之一与第一抗体结合、另一个与抗HRP抗体结合,桥抗体需用高浓度。另外,在非标记抗体酶法中可用葡萄球菌蛋白A(SPA)代替桥抗体,进行ICC染色。SPA不受种属限制,应用范围广。 (6)PAP复合物:在PAP法及酶桥法中,特别强调第一抗体和抗HRP抗体必须来自同一种属,桥抗体才能发挥桥的作用,将其连结在一起;但一些研究表明:第一抗体和抗HRP抗体来自不同种属,连结抗体仍可作为桥,将二者连在一起。例如Erlandsen发现豚鼠IgG作为第一抗体,可用羊抗兔IgG、兔PAP复合物显示之一。Grzanna(1982)根据这一报告,利用豚鼠抗DBH血清作第一抗体,羊抗兔IgG为桥抗体,12例兔血清制得的PAP复合物中,仅3例染色较佳。这种第一抗体和PAP复合物来自不同种属获得的阳性结果主要依赖于种属间的交叉反应。多数实验表明:抗体和种属交叉反应是有限的,也是不完全的。故利用桥抗体进行ICC染色时,仍以第一抗体和抗酶抗体来自同一种属为宜。 校对:2000/05/16 杨丽华 , http://www.100md.com |