|
*��ʾ����
�������ӿ�¡������һЩø
(һ)DNA�ۺ�ø
(DNA polymerase)�������ǽ�1�����������������ӵ�����(primer)��3��-OH�ϣ��ͷų�һ�����������(ppi)������ʽ��ʾ��

�ۺ�ø��Ҫ�����¼��֣�
1.�˾�DNA�ۺ�ø�� �˾�DNA�ۺ�ø(E. Coli DNA polymerase)��Ҫ��3�����ã���5����3���ľۺ����á������Ǹ���Ⱦɫ�������DNA���DNA�ϵĿ�϶�����г�RNA��������µĿ�϶����3����5��������ø���ԡ������ھۺ������в���Ĵ�������ᡣ��5����3������ø���ԡ��г������˵�DNA�������п�ƽ��(nick translation)�е�Ӧ�ã�����������ܡ�
�˾�DNA�ۺ�ø���KlenowƬ����������DNA�ۺ�ø���һ��Ƭ�Σ�ֻ����5����3�ۺ�ø���Ժ�3����5������ø���ԣ�ʧȥ��5����3����ø���ԡ����������DNA����ĩ�˳�Ϊ˫�����������32P��ǵ�����������ᣬ���ʹDNA����ͬλ�ر�ǡ����ý����и������ø�гɴ��е���ճ��ĩ�˵�DNAƬ�Σ�Ҫ�ñ��г�ƽͷĩ�˵�DNAƬ������ʱ����������KlenowƬ��ʹճ��ĩ�˵ĵ��������Ϊƽͷ��Ȼ����DNA����ø�����°�����DNAƬ������������
����д˾�DNA�ۺ�ø��ʹ˾�DNA�ۺ�ø��ǰ�߲������õ���DNA��poly(dA-dT)Ϊģ�塣����þ���Ӻ�dNTPʱ���ܱ��ֳ�ø���ԣ����Է��ϳ�DNA�����á�����û��5����3����ø���ԣ�Ҳ�����õ���DNA��Ϊģ�塣
2.�ɾ���DNA�ۺ�ø ������T4DNA�ۺ�øΪ������Ҳ����5����3�ۺ�ø���ԣ�����������ø���Աȴ˾���Ҫ��200������ˣ���Ҳ���������뵥��ĩ�˻���ͬλ�ء�
(��)RNA�ۺ�ø
RNA�ۺ�ø(RNA polymerase)��������ת¼RNA���е�RNA�ۺ�ø�бȽϸ��ӵ��ǻ��ṹ����˾�RNA�ۺ�ø������������������һ���ٽ���RNA���ӺϳɵĦ����ӣ����������ɵ��Ǧ�2�¦¦ҡ����ֽṹ��Ϊȫø(holoenzyme),��ȥ�˦����ӵ�ø��Ϊ����ø���ɾ���RNA�ۺ�ø��û���ǻ���
��������RNA�ۺ�ø�����ࡣRNA�ۺ�ø������ں����У�ת¼rRNA˳��RNA�ۺ�ø������ں����У�ת¼�����������Ҫ��TATA����RNA�ۺ�ø������ں����У�ת¼���ټ��ֻ�����tRNA������5SrRNA������Щ�ظ�˳����Alu˳�����Ҳ������øת¼�������ᵽ�ġ�TATA�����ֳ�Goldberg �CHogness˳����RNA�ۺ�ø��ĽӴ��㣬������ø��ת¼��λ�����еġ�������������ת¼�����5����һ�࣬��ת¼�������20��30��������֮����һ�θ���AT��˳������ת¼��ʼ��Ϊ0������-33��27����������-27��21������֮�䣬��һ����TATA����һ����7�������ᡣRNA�ۺ�ø��ת¼��λ�ĵ��ͽṹ��ͼ23-2��ԭ��������Ҳ���ơ�TATA����Ľṹ��RNA�ۺ�ø�����ڡ�TATAAT��(Pribnow)�к͡�TTGA-CA������

ͼ23-2 ��ת¼ø�����û���
(��)��ת¼ø
��ת¼ø(Reverse transcripatase)����RNAΪģ��ָ������������������ϳɻ���DNA( cDNA)��ø��������C�Ͳ����ķ�ת¼ø������B�Ͳ����ķ�ת¼ø����һ��������������RNA�����ķ�ת¼ø���������ǻ��ṹ�����������Ҳ����������в�ͬ�ṹ�ķ�ת¼ø������ø��Ҫþ���ӻ���������Ϊ�������ӣ�����mRNAΪģ��ʱ���Ⱥϳɵ���DNA(ssDNA)�����ڷ�ת¼ø��DNA�ۺ�ø�������£��Ե���DNAΪģ��ϳɡ����С��͵�˫��DNA(dsDNA)�����ɺ���øS1�гɶ���������˫��DNA����ˣ���ת¼ø���������κλ����mRNA��ת¼��cDNA������Ȼ��ɴ�����������������cDNA��Ҳ���������cDNA��Ϊ�����Եķ���̽�롣
(��)��������ø
�˾���������ø��(exonuclease ��)�ǴӴ�3��-OHĩ�˵�˫��DNA����3����5�������г�5���������

�˾���������ø�����Ǵӵ���DNA��3���˺�5�����г��Ѻ�����(������ʽ)��
(1)(2)

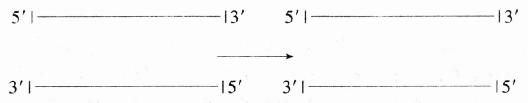
ǰһ��ø��Ҫþ�������������Ӳ��л��ԣ���һ��ø����þ���ӣ���˼�ʹ�������ϼ�EDTA�������Ҳ�л��ԡ���һ�ִ��ɾ���˸�Ⱦ�Ĵ˾�����ȡ�Ħ˺�������ø�ǴӴ���5������ĩ�˵�˫��DNA�ϣ�����г�5���������ᣬ�ڷ�Ӧʱ��Ҫþ���ӣ�
(��)����øS1
����øS1(nuclease S1)��Ҫ�ǽ��ⵥ��DNA����RNA���Ե���DNA�Ļ��Ը��ߡ�������;���г�DNAƬ�εĵ���ĩ��ʹ֮��Ϊƽͷĩ�ˣ��п��ϳ�dscDNAʱ�γɵġ����С����Լ�����DNA-RNA�Ӻ��ӵĽṹ��
(��)DNAø
DNAø��(DNase��)�����ˮ��˫������DNA��һ������ø��ʹDNA���ӽ���ɴ���5������ĩ�˵ĵ�������Ѻ�����Ļ���

�ڽ�������DNA�о�ʱ������ע���ֹDNAø����Ⱦ�������Ʊ���DNA��Ʒ���ή�⡣���������Լ����´�������Ʒ�м�EDTA������ñ������ƻ�������ø���ԡ�
(��)RNAø
RNAøA(ribonuclease A)��������ऺ������3��������ϣ��п������ں��������ӵ�5�����������һ��RNAøT1ֻ�����������ʺ������3����������п������ں��������ӵ�5�������������ķ���������Һ����Һ�ж�����RNAø������ڲ���RNA��Ʒʱ����������ף�ʵ���õIJ�������Ҫ��250��濾4h(RNAø����)������RNAø�����Ƽ�������
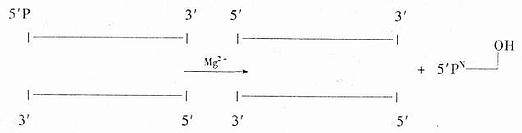
(��)����ø
��õ���T4DNA����ø(T4DNA ligase)����˫��DNA�����ڵ�3��OH��5�������֮���γ�����������������������Ӿ���ճ��ĩ�˵�����DNAƬ�Σ�����������ƽͷĩ�˵�DNAƬ�Σ�ʹ֮��Ϊһ������DNA���ӡ���������øֻ������˫��DNA���������ӵ���DNA���ӡ�T4RNA����ø����ڵ���DNA���ӻ�RNA���ӵ�5���������3��-OH֮������ɹ��ۼ���
(��)ĩ��ת��ø
ĩ������������ת��ø(terminal deoxynucleotide transferase)�������ǽ�����������ӵ�DNA���ӵ�3��-OHĩ�ˡ�
(ʮ)��������ø
��DNA��RNA���������������Ǻ��������������Ǻ������ϳ�5��-������л���һ�����;��������ø����DNA��RNA����5�����ϱ��32P���ھ�����������øø��DNAƬ�κ��ü�������ø(alkaline phosphatase)����������ֹø�е�Ƭ���������ӣ����ڿ�¡DNAƬ��ʱ�ر����á�
У�ԣ�2000/06/05 ������
, http://www.100md.com
�μ�����ҳ > ҩѧ�� > ������ѧ > ���� > ��¡
|