在ApoB100存在下,LDL受体可结合LDL;在ApoE存在下,既可结合LDL,又可结合VLDL、β-VLDL。与LDL受体不同,还有一种仅与ApoE脂蛋白结合的特异受体存在,据以下临床现象及实验结果推测还有另一种受体的存在:①纯合子FH,患者血中乳糜微粒残粒并不增加;②LDL受体缺陷的WHHL兔乳糜微粒残粒仍正常地被肝脏摄取;③LDL受体下调状态下,乳糜微粒残粒可在肝脏异化,FH的LDL受体缺陷者或WHHL兔巨噬细胞不能利用LDL使其泡沫化,但可利用含ApoE脂蛋白的乳糜微粒残粒及β-VLDL使其泡沫化,所以推测有对ApoE特异结合的第二种受体存在,即极低密度脂蛋白受体(VLDl receptor, VLDL-R)。
利用cDNA单克隆已证明有VLDL受体存在,其结构与LDL受体类似,如图6-1所示。有与LDL受体相同的五部分组成,即配体结合结构域,EGF前体结构域,含糖基结构域、跨膜结构域和胞液结构域。然而并非完全相同,配体结构域,有55%相同性(图6-4);EGF前体结构域有52%的相同性;含糖基结构域仅有19%相同性;跨膜域有32%相同性;胞浆域有46%的相同性,如图6-5、6-6所示。LDL受体对含ApoB100的LDL,含ApoE的VLDL,β-VLDL,VLDL残粒有高亲和性。VLDL受体仅对含ApoE的脂蛋白VLDL,β-VLDL和VLDL残粒有高亲和性结合并摄入细胞内,对LDL则为显著的低亲和性。VLDL受体在能量代谢活跃的心脏、肌肉、脂肪等组织细胞存在,肝脏几乎未发现,这是因为与提供组织脂肪酸机能由LPL单独承担有关。即①LPL分解结合在受体上的VLDL,水解得到的游离脂肪酸扩散通过细胞膜入细胞内,以提高利用率;②VLDL受体与LPL的mRNA有同一组织的特异性;③VLDL受体结合含ApoE的TG脂蛋白能力很强;④LPL缺损者皮下脂肪的蓄积不正常。从这几方面去考虑,VLDL受体对富含TG脂蛋白代谢起有重要作用。人VLDL受体与兔VLDL受体有97%的同源性,同人LDL受体有76%的同源性。已证实它的mRNA在组织中高度表达的结果,对这些组织细胞的脂肪酸代谢功能具有重要的意义,如肌细胞、脂肪细胞、心脏、脑和胎盘细胞等。
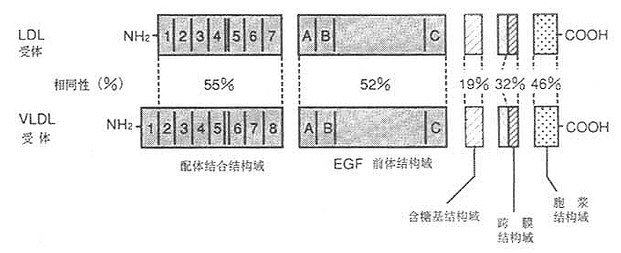
图6-5 LDL受体与VLDL受体基因结构的异同示意图

图6-6 LDL受体与VLDL受体的配体结合结构域及胞液结构域的比较
配体结构域的VLDL受体和LDL受体均保留有SDE(Ser-Asp-Glu)序列,LDL受体有7个突变序列,VLDL受体则有8个重复系列。以胞液结构域比较,LDL受体与VLDL受体均有被小窝信号肽(coated pit signal)的FDNPVY结构,其功能是与配体结合摄取进入细胞内。另外LDL受体胞液结构域,在肝细胞内侧存在向基运输小窝(basipetal translocation pit)的[RNxDxx(S/T)xxS]结构,VLDL受体则无此小窝,如图6-6所示。
LDL受体受细胞内胆固醇负反馈抑制,VLDL受体则不受其负反馈抑制,当VLDL受体的mRNA量成倍增加时,不受LDL乃至β-VLDL的影响。这是因为VLDL的配体关系使β-VLDL的摄取不受限制。这一点,对由单核细胞由来的巨噬细胞的泡沫化在早期动脉粥样硬化的斑块形成中有重要意义。
VLDL受体在脂肪细胞中多见,可能与肥胖成因有关。
, http://www.100md.com