-第一节 胆固醇酯转移蛋白
脂类代谢过程中的非极性CE和TG以及载脂蛋白,在脂蛋白之间进行转运和交换。这种交换转运是否需要载体,如何转运,各脂蛋白之间的脂质和蛋白又如何达到平衡,早在60年代,人们已开始了饶有兴趣的研究。1968年Akenumz与Glomset等发现血浆中有LCAT能催化HDL上Ch变成CE,转运到CM及VLDL或其他脂蛋白上,并很快达到平衡。其后陆续有多种参与脂蛋白代谢的蛋白质因子被发现。从蛋白质的生物学性质及功能看,这类蛋白因子似乎是酶,但又不具备酶学特征;似乎是受体,也不具备作为受体的特点,这种似酶非酶,是受体非受体的蛋白质,本章只好暂且称其为脂类代谢有关的特殊蛋白质。
第一节 胆固醇酯转移蛋白
早在1975年,Zilvermit等发现兔血浆无脂蛋白部分含有使胆固醇酯转运的特殊蛋白质。Barter等报道兔血浆d>1.21g/ml部分存在有促进脂蛋白和TG交换的蛋白因子,其后从高胆固醇血症兔血浆中分离得到这种活性蛋白,能促使HDL与LDL和VLDL之间进行CE交换,以后的研究中又发现这种质白质与动脉粥样硬化发生密切相关,这种特殊蛋白被之谓胆固醇酯转移蛋白(cholester ester transfer protein, CETP)。
一、CETP结构
CETP又称为脂质转运蛋白(lipid transfer protein, LTP),从血浆d>1.21g/ml组份中精制得到,是由467个氨基酸残基组成的单链多肽,其中非极性氨基酸残基高达45%,是一种疏水性蛋白质,很容易被氧化而失活。CETP由肝脏、小肠、肾上腺、脾脏、脂肪组织及巨噬细胞合成,细胞内成熟蛋白分子量为74000。最近已阐明其结构编码基因存在于第16染色体。与LCAT的基因靠近。CETP基因由16个外显子和15个内含子约20.5kb组成。
二、CETP生理功能
CETP促进各脂蛋白之间脂质的交换和转运。他催化HDL3上的胆固醇酯转运到富含ApoB的脂蛋白VLDL和LDL上,其中的TG经HTGL作用水解,使HDL颗粒缩小,CETP在完成和促进胆固醇逆转运过程中充当重要角色。周围组织细胞膜的游离胆固醇与HDL结合后,被LCAT酯化成胆固醇酯,移入HDL核心,并可通过CETP转移给VLDL,LDL,再被肝脏的LDL及VLDL受体摄取入肝细胞,至此,完成了胆固醇从周围末梢组织细胞经HDL转运到肝细胞的过程,称之为胆固醇的逆转运,如图7-1所示。
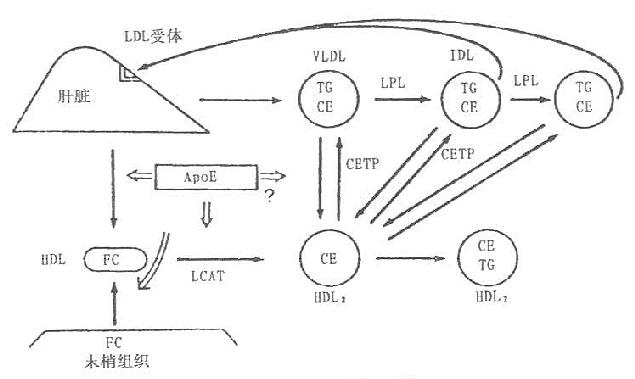
图7-1 胆固醇逆转运系统
目前认为,血浆中各脂蛋白的胆固醇酯主要通过LCAT和CETP的共同作用生成。CETP与LCAT一样也常与HDL结合在一起。
三、CETP缺乏症
血浆中CETP缺乏,HDL中CE蓄积TG降低,无法转运给VLDL及LDL,出现高HDL血症,从而使VLDL、LDL的CE减少及TG增加。这是因为从HDL将CE转运给含ApoB脂蛋白的途径出现障碍所致。利用酶联免疫方法检测血浆中CETP活性,其活性很低。
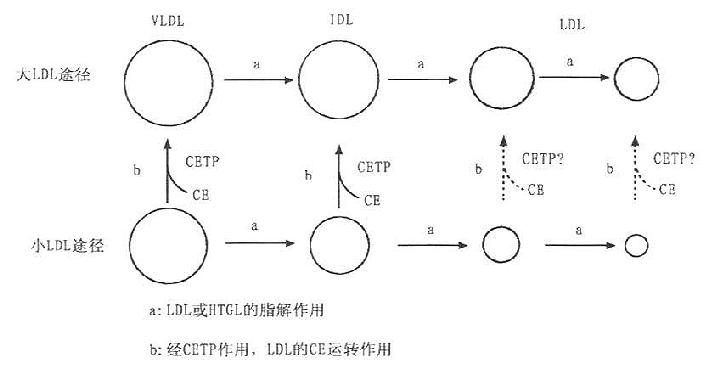
图7-2 LDL形成的两条途径
因CETP的缺陷可引起高HDL血症,其中出现有富含ApoE和CE的类似于HDLc的多相分散LDL(polydisperse LDL)。利用平衡密度梯度超速离心可使这种多相分散LDL显示出16个亚组份。采用聚丙烯酰胺梯度凝胶电泳,血胆固醇水平正常的人LDL仅显示一个单一的区带。若CETP缺乏者,LDL可显示两条区带,其后进一步阐明,这两个区带是属于密度相同,其理化性质有差异的大小两种颗粒的LDL。究其原因,是因为LDL的成熟过程有大LDL和小LDL途径,如图7-5所示。ECTP的作用是促使HDL的CE转运到LDL中,使其形成成熟均一的LDL,CETP是LDL成熟的必]需因素。
利用酶联免疫法测定CETP缺陷者,血清CETP水平低于健康人,其量与HDL-C呈负相关。实验证明,CETP缺陷的纯合子,转运CE的作用完全丧失。经PCR及DNA测序确认,CETP缺陷常在基因内含子14的剪接供体位点上易发生G→A突变。据报道,CETP缺陷者可出现于杂合子,又可出现于纯合子。据此推测,CETP缺陷者可能是一种复合型异型合子(compound heterozygote)。
CETP缺陷是引起高HDL血症的原因,然而血浆中CETP的活性差异很大。血浆CETP活性低的动物,胆固醇负荷实验发现,很易引起动脉粥样硬化,而HDL-C呈高值;CETP活性高的动物进行胆固醇负荷实验发现,也很容易引起动脉粥样硬化,然而HDL-C为低值。这种有趣的现象令人费解。然而最大的可能是,CETP以几种不同的表型发挥着不同的生理功用之故。因此,与其说CETP的某一表型是CE转运蛋白还不如说是致动脉粥样硬化的因子。
总之,CETP缺陷是引起动脉粥样硬化的重要因素之一。
, 百拇医药