 |
 |
关键词:成纤维细胞;维甲类X受体;维甲酸反应元件;维甲酸
摘要 目的: 确定人真皮成纤维细胞中维甲类X受体(RXR)与维甲酸反应元件(β2 RARE)的结合。方法: 培养成纤维细胞,反式维甲酸(At-RA)刺激细胞不同时段,抽提核蛋白,32 P标记β2 RARE作为探针,并加入RXRα和β抗体,进行凝胶阻滞分析。结果: 对照组细胞核蛋白与β2 RARE形成复合体Ⅰ;At-RA刺激细胞后,迅速诱导产生明显的复合体Ⅱ。选择两组核蛋白(对照组和At-RA刺激4h组),分别加入RXRα和β抗体,凝胶阻滞分析发现RXRβ抗体可使这两组核蛋白产生新的上移复合体SS,RXRα抗体则无此作用。结论: RXRβ能与β2 RARE结合,它们的结合不受AT-RA的影响;提示RXRβ可能是作为一辅助因子参与了RARβ2 的转录调节。
中国图书资料分类法分类号 Q753
STUDY ON THE BINDING OFRETINOID X RECEPTOR WITH β 2 RARE
Xie Xiaoxun
( Department of Histology & Embryology,Guangxi Medical University,Nanning 530021)
Abstract Objective: Toconfirm the binding of Retinoid X Receptors(RXR) with Retinoic Acid Responsive Element (β2 RARE).Method: Fibroblastsin human derma were cultured with stimulation for All-trans Retinoic Acid (At-RA).Labelingβ2 RARE with 32 P as a probe,nuclear extract was isolated from thesecells and incubated with antibody to RXR α and β,then gel retardation assays wereperformed.Results: The nuclear protein from control group was binded withβ2 RARE to form a complex Ⅰ.Stimulating cells with At-RA quickly induced anobvious complex Ⅱ.Two groups of nuclear protein (control group and the group of 4 hstimulated with At-RA) were selected for incubating separately with antibody to RXR α andβ.The gel retardation assays showed a new supershift complex SS after using antibody toRXR β for incubating with the nuclear proteins of these two groups,while the antibody toRXR α did not appear to have this effect.Conclusion: RXR β binded to β2 RARE.AT-RA has no effect on this binding.It suggested that RXR β may be an accessoryfactor in transcriptional regulation of RARβ2 .
Key words fibroblast;retinoid X receptors (RXR);retinoic acidresponsive element (RARE);retinoic acid (RA)
维甲酸(Retinoic Acid,RA)为维生素A的衍生物,其信号转导通路是由细胞内RA结合蛋白、维甲酸受体(RetinoicAcid Receptor,RAR)、维甲类X受体(Retinoid X Receptor,RXR)、维甲酸反应元件(RetinoicAcid Responsive Element,RARE)和靶基因组成,具有传递RA信号,控制基因表达以及参与脊椎动物发育、细胞分化和维持体内平衡的作用。RA进入靶细胞后,由RA结合蛋白运送到细胞核,与核受体RAR和RXR结合,识别DNA上的维甲酸反应元件,通过反式作用调控靶基因[1] 。β2 RARE为一种维甲酸反应元件,目前认为能与RAR和RXR等一些受体结合,起转录调节作用[2] 。由于RAR和RXR中有众多亚型及异构体,而且在不同的组织及细胞中表达亦不相同。因此,具体哪一种核受体与β2 RARE结合,目前尚未完全清楚。本研究通过体外培养人真皮成纤维细胞,抽提核蛋白,标记β2 RARE作为探针,与核蛋白结合,进行凝胶阻滞分析(GelRetardation Assay),并在此基础上进行抗体结合凝胶阻滞分析,旨在证明β2 RARE能否与RXR结合,与哪种RXR结合,使我们对复杂的RA信号转导通路有进一步的认识。
1 材料和方法
1.1 细胞培养:将常规手术切除的包皮,浸泡于0.25%trypsin液中,4℃过夜;机械分离上皮和真皮层,绞碎真皮,10g/L胶原蛋白酶孵育3h,37℃。将成纤维细胞转入含10%小牛血清的DMEM液中培养,5%CO2 ,37℃。饥饿细胞48h(即生长在含0.5%小牛血清的DMEM液中),然后用1mmol/L全反式维甲酸(AT-RA,Sigma产品)刺激细胞不同时段。避光条件下进行稀释RA,每次均为新鲜制备。
1.2 核蛋白制备[2] :对照组细胞用含10%小牛血清的DMEM培养。实验组细胞用0.5%小牛血清的DMEM培养48h后,用RA刺激不同时段。4℃PBS冲洗细胞,冰上收集细胞,离心后用细胞体积5倍的BufferA(含10 mmol/L Hepes pH7.9,1.5 mmol/L KCl,2 mmol/L DTT)将细胞打散,再离心去上清液,用细胞体积3倍的BufferA将细胞悬浮,冰育10min后,用均化器冲击10次,使细胞破裂,离心去上清液,收集细胞核。用细胞核总体积3/4倍的BufferC(含20 mmol/L Hepes pH7.9,20% glycerol,0.5 mmol/L NaCl,1.5 mmol/L MgCl2 ,0.2 mmol/L EDTA,2 mmol/L DTT)将细胞核悬浮,置于冰上30min,期间每5 min振震一次,使细胞核破裂并在显微镜下观察细胞核裂解情况,随后离心收集含核蛋白的上清液;并用BufferD(20 mmol/L Hepes pH7.9,20% glycerol,100 mmol/L KCl,0.2 mmol/L EDTA,1.5 mmol/L MgCl2 ,0.2 mmol/L DTT)进行透析,4℃,4h。确定蛋白质浓度(Protein test kit,Bio-Rad),-70℃贮存备用。
1.3 探针制备:合成β2 RARE寡核苷酸(GGGTAGGGT-
TCACCGAAATTCACTCG),用32 P-rATP进行末端标记(约1×105 cpm)。
1.4 凝胶阻滞分析(Gel Retardation Assay)
1.4.1 核蛋白与探针的结合反应:取3~5 μg核蛋白,1μg poly dI-dc,0.5~1ng32 P标记的β2 RARE至结合缓冲液(10 mmol/L EDTA,2.5 mmol/L MgCl2 ,0.2 mmol/L DTT);探针-核蛋白结合反应液总体积为25μl,室温下孵育20min,加入1μl电泳指示液(50%甘油:0.1%溴酚蓝=2∶1)。
1.4.2 探针-核蛋白复合物的凝胶阻滞分析:将探针-核蛋白复合物进行5%聚苯稀酰胺凝胶电泳,1~1.5h;电泳完毕将凝胶翻转置滤纸上,真空干燥,65℃,1h。柯达胶片于-70℃进行放射自显影。
1.4.3 组间对照组的设置:为了保证探针与核蛋白结合的特异性,避免由同位素非特异性的结合引起的假阳性,在结合反应液中预先加入未标记探针(约为实验组量的100倍),室温下孵育20min,然后再加入标记探针进行凝胶电泳,其余步骤同上。
1.4.4 抗体-核蛋白-探针结合反应及凝胶阻滞分析:分别加入2μlRXRβ抗体(Affinity Bioreagent公司产品)和RXRα抗体(Santa Cruz Biotechnology公司产品)与核蛋白孵育,4℃,16h。然后按上述步骤与标记探针结合并进行凝胶阻滞分析。
2 结 果
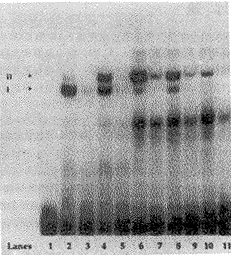
2.1 未经RA刺激的对照组,显示有核蛋白与β2 RARE的结合,形成明显的复合体Ⅰ,和隐约可见的复合体Ⅱ(图1,第2泳道)。RA刺激细胞1h的实验组,迅速诱导产生明显的复合体Ⅱ,经密度测量仪测量,复合体Ⅰ、Ⅱ的密度基本相等(图1,第4泳道)。RA刺激细胞4h的实验组,复合体Ⅱ明显增强,复合体Ⅱ的密度为复合体Ⅰ的10倍(图1,第6泳道)。此后,随着RA刺激时间的延长,复合体Ⅰ的密度减弱并消失;同时复合体Ⅱ亦逐渐减弱(图1,第8、10泳道)。预先加入过量的未标记探针与核蛋白孵育,再用标记的探针进行检测,复合体基本消失(图1,第3、5、7、9、11泳道),说明复合物的形成不是由同位素非特异性结合所致,而是β2 RARE与核蛋白特异结合的结果,虽然部分复合体未完全消失(图1,第7、9泳道),可能与加入的未标记探针量不足有关。
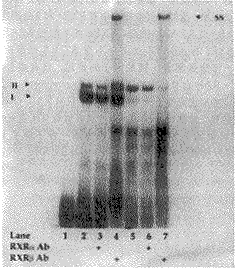
图1 β2 RARE-核蛋白结合的凝胶阻滞分析图谱
空白对照(第1泳道);未经RA刺激的对照组(第2泳道);RA刺激1,4,6,和24h组的核蛋白(第4,6,8,10泳道),显示核蛋白与β2 RARE结合形成复合体Ⅰ,Ⅱ;加入100倍过量的未标记探针的对照组(第3,5,7,9,11泳道),显示复合体Ⅰ,Ⅱ基本消失
2.2 为了确定β2 RARE-核蛋白复合体中含有何种RXR受体,分别选用了RXRα和β抗体,与RA刺激4h的实验组和无RA刺激组(对照组)的两组核蛋白孵育,结果为加入RXRβ抗体的两组核蛋白均产生新的上移复合体SS(图2,第4,7泳道);与此相反,加入RXRα抗体则无上移复合体SS的出现(图2,第3,6泳道)。
图2 RXR抗体-核蛋白-β2 RARE结合的凝胶阻滞分析图谱
空白对照组(第1泳道),未经RA刺激的对照组(第2,3,4泳道);RA刺激4h组(第5,6,7泳道);加入RXRα抗体组(第3,6泳道);加入RXRβ抗体组(第4,7泳道)出现新的上移复合体SS
3 讨 论
RA的生物学作用是通过核受体RAR和RXR介导,这些受体均有不同的亚型(α、β、γ),每一亚型中又存在着多种异构体。RA与这些受体结合后,不但可与靶基因上的反应元件结合,调控靶基因的转录,而且还可以和这些受体异构体上的反应元件结合,调节自身受体的表达[3] ,如RARβ的异构体RARβ2 ,其启动子区域上含有反应元件β2 RARE,目前认为RA与RAR或RXR结合后,可识别β2 RARE并与之结合,调节RARβ2 的表达。有意义的是,现发现在一些肿瘤细胞株中,缺乏RARβ2 的表达,当将RARβ2 基因转入这些肿瘤细胞后,肿瘤细胞的生长受抑制;虽然RARβ2 的这一作用与肿瘤抑制基因P53 相似[4~8] ,但其机制尚不太清楚。因此,明确β2 RARE与何种维甲酸受体结合,有助于进一步认识RARβ2 在肿瘤生长抑制中的作用机制。
从本研究结果来看,在无RA刺激的情况下,可见明显的复合体Ⅰ和隐约的复合体Ⅱ,说明细胞核中含有能与β2 RARE结合的蛋白。在给予RA刺激细胞1h后,复合体Ⅱ迅速增强,至RA刺激4 h时最为明显;已有研究表明[9] ,RA刺激人真皮成纤维细胞后,可使RARβ和γ的信息RNA表达迅速增强,由此提示本研究结果中复合体Ⅱ的出现,可能与RARβ和γ被迅速诱导产生有关。与复合体Ⅱ不同,复合体Ⅰ与RA的作用无明显关联,这与真皮成纤维细胞中只有RXRα和β的表达,且不受AT-RA影响的报道相符[10] ,因此推测这些复合体中有可能存在着RXRα和β。为了进一步弄清RXR与β2 RARE结合的情况,我们通过抗体结合凝胶阻滞分析方法,发现只有RXRβ抗体能使这两组核蛋白产生新的复合体SS,说明这两组核蛋白中含有特异性蛋白RXRβ,RXRβ与相应抗体结合后,再与β2 RARE结合形成较大的分子,使电泳上呈现移动缓慢的条带,即复合体SS。这一结果说明,虽然成纤维细胞中有RXRα和β的表达,但只有RXRβ与β2 RARE结合,RXRα并不参与β2 RARE的结合;RXRβ与β2 RARE的结合不受AT-RA的影响,提示RXRβ可能是起着一辅助因子的作用。已有体外实验报道[11] ,RAR自身形成的同二聚体(homodimer),因不能识别相应的反应元件而无生物学活性,只有RAR-RXR结合形成的异二聚体(hereodimer),才能有效地与反应元件结合,发挥转录调节作用。本研究作为第一步了解了RXR与β2 RARE的结合情况,但有关RAR与β2 RARE的结合尚不清楚,有待于用该方法作进一步的检测。
(本实验主要部分在美国哥伦比亚大学医学院中心实验课完成,Dr.MPeacoke给予指导,特此感谢)
参考文献
1 Gigure V.Retinoic acid receptor andcellular retinoid binding pro-tein:interplay in retinoid signaling.Endocrine Review,1994,15(1):61
2 Lee X,Si SP,Peacocke M.Cellular aging and transformation suppression:a role for re-tinoic acid beta 2.Exp.Cell Res,1995,218(2):296
3 de Th'ee H,Vivano Ruiz MM,Tiollais P,et al.Identification of a retinoic acidresponsive element in the retinoic acid receptor beta gene.Nature,1990,343:177
4 Swisshelm K,Ryan K,Lee X,et al.Down-regulation of retinoic accid receptor beta inmammary carcinoma cell line and its up- regulation in senescing normal mammary epithelialcells.Cell Growth Differ,1994,5:133
5 Hu L,Crowe,DL,Rheinwarld JG,et al.Abnormal expression of retinoic acid receptors andkeratin 19 human oral and epidermal squaous cell line.Cancer Res,1991,51:3972
6 Houle B,Leduc F,Bradley WE.Implication of RARβ in epi-dermoid (squamous) lungcancer.Genes.Chromosomes Cancer,1991,3:358
7 Gebert JF,Moghal N,Frangioni JF,et al.High frequency of retinoic acid receptor betaabnormalities in human lung cancer.Oncogene,1991,6:1 859
8 Si SP,Tsou HC,Lee X,et al. RARβ2 -Mediated Growth inhibition in HeLacells.Exp.Cell Res,1996,223:102
9 Tsou HC,Lee X,Si SP,et al.Regulation of retinoic acid expression in dermalfibroblast.Exp.Cell Res,1994,211:74
10 谢小薰,Tsou HC,Peacocke M.全反式维甲酸对人真皮成纤维细胞RXR的表达影响.暨南大学学报,1998,19(6):68
11 Mangelsdorf DJ,Umesono K,Kliewer SA,et al.A direct repeat in the cellular retinol-binding protein type Ⅱ gene confers differential regulation by RXR andRAR.Cell,1991,66:555
收稿日期:1999-01-10, http://www.100md.com