血小板衍生生长因子与肝纤维化的研究进展
http://www.100md.com
王萍. 袁农.
血小板衍生生长因子.肝纤维化.抗纤维化.
参见附件(180kb)。
作者:王萍 袁农
单位:(陕西省人民医院消化研究室,西安 710068)
关键词:血小板衍生生长因子;肝纤维化;抗纤维化
西安医科大学学报000344摘 要:肝纤维化是一种常见的慢性进行性肝病。血小板衍生生长因子(PDGF)为重要的促细胞活化、分裂、增殖剂,在肝纤维化的发生、发展中起着非常重要的作用。PDGF与其相应受体结合后经细胞信号传导才能发挥作用,晚近通过对PDGF与其受体结合后细胞内信号传导途径的研究,发现阻断其信号传导有可能阻断PDGF的作用,发挥抗肝纤维化作用。
中图分类号:R575.2 文献标识码:A
文章编号:0258-0659(2000)03-0293-03
肝纤维化是一种常见的慢性进行性肝病。肝脏星形细胞(Hepatic stellate cell ,HSC)的激活、增殖以及细胞外基质的沉积是肝纤维化的两个重要环节[1]。血小板衍生生长因子(Platelet-derived growth factor,PDGF)能够强烈刺激肝星形细胞的增殖、迁移,促使其胶原的产生和沉积,在肝纤维化发生发展中起着非常重要的作用[2]。因此抑制PDGF的产生或对其作用进行拮抗是抗肝纤维化的重要途径。
1 PDGF及其受体的分类及结构
1.1 PDGF分类、结构及来源
PDGF可分为分子量为31KD含有7%糖的PDGF I及28KD含4%糖的PDGF Ⅱ。两者均由两条高度同源的A链及B链组成,这使PDGF具有三种形式的二聚体结构,即PDGF-AA,PDGF-BB及PDGF-AB[2,3]。
体内单核/巨噬细胞是主要合成PDGF的细胞。在生理状态下,PDGF以α颗粒的形式储存于血小板中,肝脏受损时,巨噬细胞、血小板、浸润的炎细胞、受损的内皮细胞及激活的肝星形细胞均可以分泌PDGF。以自分泌、旁分泌的方式发挥作用[4,5]。
1.2 PDGF受体分类、结构及激活方式
PDGF必须与细胞膜上的相应受体结合后才能发挥其生物学效应。PDGF受体由两种亚单位α及β构成,其分子量为170~180KD。二者与PDGF结合力相差很大,α单位与PDGF A链及B链有较高的亲和力,而β亚单位仅与B链有高亲和力。所以α亚单位可与PDGF-AA、PDGF-AB及PDGF-BB结合,β亚单位仅与PDGF-BB及PDGF-AB结合[2,3]。Wang等[5]人的研究证明,在肝纤维化时,HSC表面的PDGF受体以β受体为主,与PDGF-BB具有较强的亲和力,认为PDGF-BB以及PDGF-β在肝纤维化过程中的作用尤为突出。
PDGF受体是一种跨膜糖蛋白,具有酪氨酸蛋白激酶活性,由细胞外N端与PDGF特异识别的结构域、单链顺序跨膜的中间疏水结构域和细胞内C端具有酪氨酸蛋白激酶活性的肽段结构域组成。当受体与其配体结合后促使两个受体分子形成二聚体,激活细胞内结构域酪氨酸残基自身磷酸化,或促使激活特殊靶蛋白的酪氨酸残基磷酸化,从而将信号传入细胞内,经级联式放大瀑布效应调控细胞的生命活动,包括靶细胞的分裂增殖[6]。
2 PDGF生物学效应和在肝纤维化中的作用
实验证明PDGF是一种重要的促有丝分裂因子,具有刺激特定细胞群分裂增殖的能力。肝受损时,大量分泌的PDGF刺激间质星形细胞增殖,转化为肌纤维样母细胞,并促使星形细胞迁移,聚集于炎症受损区[7]。而肌纤维样母细胞合成大量细胞外基质沉积于肝细胞胞间质,促进肝纤维化发生。
PDGF能够促进肌纤维母细胞产生胶原,尤其是I型及Ⅲ型胶原。Isbrucker[1]在体外培养的实验中证明,PDGF可促使未融合猪肝肌纤维样母细胞增殖,使已融合的细胞产生胶原,但对其细胞数量无影响。
PDGF还可通过上调组织金属蛋白酶抑制剂(TIMP-1)抑制胶原酶的作用,以减少细胞外基质的降解[4,8]。
3 PDGF与抗肝纤维化
3.1 PDGF的细胞内信号传导
抑制PDGF的产生和阻断其促纤维化的作用可望成为抗肝纤维化的途径,晚近通过对PDGF与其受体结合后细胞内信号传导途径的研究,发现阻断其信号传导能阻断PDGF的作用[9]。
PDGF与其受体结合后,可激活细胞内酪氨酸蛋白激酶(TPK),PDGF受体发生自动磷酸化,受体位点上的磷酸化酪因同源区2(Src-SH2)或酪氨酸磷酸区域进行结合[10]。结合于已激活的PDGF受体的分子包括有磷酯酰肌醇三激酶(PI 3-K),细胞外信号调节激酶(ERK)、磷酯酶C、酪氨酸激酶及Crb2、shc、shb等。经过一系列磷酸化反应,可活化核膜上丝氨酸(Ser)或苏氨酸(Thr)残基激酶,后作用于核内蛋白转录因子,如癌基因fos、jun、myc等编码的蛋白质,经直线传递信号使DNA的一些活性转录区参与基因表达的调控,或增加拓扑异构酶活力,改变细胞RNA含量,使细胞发生一系列表型改变,促细胞生长和分裂[7]。(如图1)。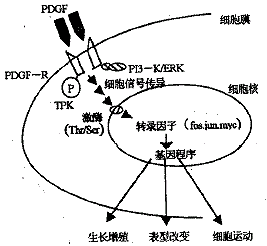
图1 PDGF的细胞内信号传导途径
在PDGF发挥的作用中,PI 3-K及ERK途径是膜内信号传导的两个最重要途径。同时,PDGF引起细胞[Ca2+]浓度增加也促进了细胞增殖[6]。
3.2 细胞外信号调节激酶(ERK)及其抑制剂
业经证明ERK激活对激发细胞增殖是必不可少的。同时ERK途径与细胞外基质成分胶原的产生有密切关系[11]。ERK有两种构型,即ERK1(P44MAPK)和ERK2(P42MAPK)。人类肝星形细胞在ERK激活后继有c-fos激活,进一步作用于DNA转录因子,促细胞分裂[12]。故对ERK的作用进行干预抑制PDGF诱导的HSC增殖。若预先将HSC与升高细胞内cAMP水平的药物孵育,能明显降低PDGF诱导的ERK磷酸化、ERK活性、c-fos表达及促有丝分裂作用。己酮可可碱(Pentoxiffilly, PTF)是一种磷酸二酯酶抑制剂,可促细胞内cAMP浓度升高,抑制ERK传导途径和细胞增殖[13]。Preaux[14]实验证明,PTF可以减少PDGF诱导的细胞增殖,抑制Ⅰ、Ⅲ型胶原的合成,并使Ⅰ、Ⅲ型胶原mRNA减少,说明抑制胶原合成发生在转录前水平。
3.3 磷酯酰醇三激酶(PI 3-K)及其抑制剂
PI 3-K聚集激活PDGF受体,它由85KD的调节亚单位(2个SH-2区域)和110KD的催化亚单位构成。PDGF刺激PI 3-K与其受体结合,使85KD亚单位酪氨酸磷酸化后,激活蛋白激酶C和蛋白激酶B。这一信号传导引起细胞增殖及迁移,并聚集于炎症区[15]。PDGF-AA、AB及BB均可激活PI 3-K,以PDGF-BB作用最强,这与其促细胞分裂增殖能力有关。PDGF-BB对HSC的增殖和化学趋化性起作 用,而PDGF-A只促HSC增殖,无化学趋化作用,这可能与结合于不同受体,以及与受体后信号传导途径不同有关。渥曼青霉素(wortmannin)是一种真菌代谢产物,可结合PI 3-K,非竞争性抑制PI 3-K活性;但对受体的自动磷酸化和85KD蛋白与受体之间的联系无影响。渥曼青霉素还能部分抑制ERK活性,对它的深入研究,有可能成为一种新的抗纤维化药物[7]。
3.4 钙离子及其抑制剂
钙离子浓度的变化也与细胞增殖有关。实验证明PDGF明显升高细胞内[Ca2+][17]。其机理可能为PDGF与其受体结合后,酪氨酸激酶自动磷酸化,诱导磷酸肌醇(IP)循环,其循环产生IP3作用于细胞内皮网特异受体使其Ca2+释放,引起细胞内Ca2+水平迅速增加启动细胞内信号系统(I相)。IP3进一步磷酸化产生IP4又促使细胞外Ca2+内流 ,即细胞内Ca2+浓度的慢反应(Ⅱ相)[16]。证实PDGF诱导的Ca2+升高主要为细胞外Ca2+内流,与“T”型通道打开有关[17]。发现有机物氯化镍抑制“T”型通道,故有可能抑制PDGF所诱导的有丝分裂。因此,开发“T”通道特异抑制剂可阻断肝纤维化[9]。国内学者[18]选用细胞外钙离子拮抗剂维拉帕米(ver)和细胞内钙离子拮抗剂尼卡地平(nic),观察其对大鼠肝细胞胶原合成及升钙作用,结果显示ver可抑制PDGF促胶原合成和升钙效用,若ver与nic合用效果更佳。
3.5 细胞Na+-H+交换途径及其抑制
Di Sario[19]等人的研究表明:HSC激活伴随着Na+-H+交换的激活,细胞内pH增加。这种激活依赖于钙离子-钙调蛋白及蛋白激酶C途径。证明PDGF可增加Na+-H+交换活性,促进细胞分裂。Na+-H+交换抑制剂阿米洛利(amiloride)可抑制PDGF诱导的细胞增殖,故可能有抗纤维化作用。
3.6 酪氨酸蛋白激酶(TPK)及其抑制剂
PDGF与受体结合,激活受体细胞膜内部分的TPK,引起一系列信号传导及调节细胞增殖。因此阻断PDGF作用的关键之一是阻断TPK的活性。槲皮素(quercetin)是一种TPK抑制剂,属黄酮类,可以阻断细胞膜内酪氨酪残基的磷酸化使TPK失活,PDGF不能发挥作用[20]。故对TPK抑制剂的开发可望成为阻断纤维化的另一途径。
参考文献:
[1]Isbrucker RA,Oeferson Tc.Platelet-derived growth factor and pentoxifylline modulation of collagen synthesis in myofibroblasts.Toxicology and Applied[J].Pharmacology,1998,149(1)∶120
[2] Ikura Y,Morimoto H,Ogami M ,et al.Expression of platelet-derived growth factor and its receptor in livers of patients with chronic liver disease[J].J Castroentrology,1997,32(4)∶496
[3] Pinani M,Milani S, Herbst H ,et al.Expression of platelet-derived growth factor and its receptors in normal human liver and during active hepatic fibrosis[J].Am J Pathol,1996,148(3)∶785
[4] Burt AD,Cellular and molecular aspects of hepatic fibrosis[J].J Pathology,1993,170(2)∶105
[5] Wang L,Yamasaki G, Johnso R ,et al.Induction of β-platelet-derived growth factor receptor in rat hepatic lipocytes during cellular activation in vivo and in culture[J].J Clin Invest,1994,94(4)∶1563
[6] Friedman SL.Closing in on the signals of hepatic fibrosis[J].Grastroenterology,1997,112(4)∶1406
[7]Marra F,Gentilini A,Pinzani M,et al.Phosphatidylinositol 3-kinase is requried for plateled-derived growth factor′s actions on hepatic stellate cells[J].Gastroenterology,1997,112(4)∶1297
[8] Herbst H ,Schupan D,Milani S.Fibrogenesis and fibrolysis in ths liver[J].Verh Dtsch Ges Pathol,1995,79(1)∶15
[9] Pinzanj M,Marra F, Carlonj V.Signal transduction in hepatic stellate cells[J].Liver,1998,18(1)∶2
[10] Pawso T, Protein modules and signalling networks[J].Nature, 1995,373(4)∶573
[11] Davis BH,Chen A,Beno DWA.Raf and mitogen-activated protein kinase regulate stellate cell collagen gene expression[J].J Biol Chem,1996,271(3)∶11039
[12]Marra F,Ghosh choudhury G,Abboud HE.Interferon-gamma mediated activation of statl α regulates growth factor induced mitogenesis[J].J Clin Invest,1996,98(3)∶1218
[13] Pinznj M,Marra F,Gallgiuki A ,et al.Inhibiton by Pentoxifylline of extracelluar signal-regulated kinase activaton by plateletderived growth factor in hepatic stellate cells[J].Br J Pharmacol,1996,119(6)∶1117
[14] Preaux AM, Mallat A, Rosenbaum J,et al.Pentoxiffyline lnhibits growth and collagen synthesis of cultured human hepatic myofibrobloast-like cells[J].Hepatology,1997,26(2)∶315
[15] Valius M,Kazlaukas A.Phospholipase C-gamma and phosphatidylinostiol 3-kinase are the downstream mediators of the PDGF receptor′s mitogenic signal[J].Cell,1993,73(1)∶321
[16] Failli P,Ruocco C, Franco RD ,et al.The mitogenic effect of platelet-derived growth factor in human hepatic stellate cells requires calcium influx[J].Am J Physiol,1995,269(38)∶C1133
[17] Wang Z, Estacion M, Mordan LJ.Ca2+ Influx via-T type channels modulates PDGF-Induced replication of mouse fibroblasts[J].Am J Physiol,1993,265(39)∶C1239
[18] 姚迪,李定国,徐芹芳.血小板衍生生长因子和钙离子拮抗剂对大鼠肝细胞胶原合成及细胞内游离钙浓度的影响[J].胃肠病学和肝病学杂志,1998,7(2)∶132
[19] Di Sario A,Sregliati Baroni G, Bendia E, et al.Characterization of ion transport mechanisms regulating intracellular pH in hepatic stellate cells[J].Am J Physiol ,1997,273(36)∶G39
[20] Levitzki A, Gazit A.Tytosine kinase inhibition:an approach to drug development[J].Science, 1995,26T(5)∶1782
(收稿日期:1999-12-08 修回日期:2000-01-10)
作者:王萍 袁农
单位:(陕西省人民医院消化研究室,西安 710068)
关键词:血小板衍生生长因子;肝纤维化;抗纤维化
西安医科大学学报000344摘 要:肝纤维化是一种常见的慢性进行性肝病。血小板衍生生长因子(PDGF)为重要的促细胞活化、分裂、增殖剂,在肝纤维化的发生、发展中起着非常重要的作用。PDGF与其相应受体结合后经细胞信号传导才能发挥作用,晚近通过对PDGF与其受体结合后细胞内信号传导途径的研究,发现阻断其信号传导有可能阻断PDGF的作用,发挥抗肝纤维化作用。
中图分类号:R575.2 文献标识码:A
文章编号:0258-0659(2000)03-0293-03
肝纤维化是一种常见的慢性进行性肝病。肝脏星形细胞(Hepatic stellate cell ,HSC)的激活、增殖以及细胞外基质的沉积是肝纤维化的两个重要环节[1]。血小板衍生生长因子(Platelet-derived growth factor,PDGF)能够强烈刺激肝星形细胞的增殖、迁移,促使其胶原的产生和沉积,在肝纤维化发生发展中起着非常重要的作用[2]。因此抑制PDGF的产生或对其作用进行拮抗是抗肝纤维化的重要途径。
1 PDGF及其受体的分类及结构
1.1 PDGF分类、结构及来源
PDGF可分为分子量为31KD含有7%糖的PDGF I及28KD含4%糖的PDGF Ⅱ。两者均由两条高度同源的A链及B链组成,这使PDGF具有三种形式的二聚体结构,即PDGF-AA,PDGF-BB及PDGF-AB[2,3]。
体内单核/巨噬细胞是主要合成PDGF的细胞。在生理状态下,PDGF以α颗粒的形式储存于血小板中,肝脏受损时,巨噬细胞、血小板、浸润的炎细胞、受损的内皮细胞及激活的肝星形细胞均可以分泌PDGF。以自分泌、旁分泌的方式发挥作用[4,5]。
1.2 PDGF受体分类、结构及激活方式
PDGF必须与细胞膜上的相应受体结合后才能发挥其生物学效应。PDGF受体由两种亚单位α及β构成,其分子量为170~180KD。二者与PDGF结合力相差很大,α单位与PDGF A链及B链有较高的亲和力,而β亚单位仅与B链有高亲和力。所以α亚单位可与PDGF-AA、PDGF-AB及PDGF-BB结合,β亚单位仅与PDGF-BB及PDGF-AB结合[2,3]。Wang等[5]人的研究证明,在肝纤维化时,HSC表面的PDGF受体以β受体为主,与PDGF-BB具有较强的亲和力,认为PDGF-BB以及PDGF-β在肝纤维化过程中的作用尤为突出。
PDGF受体是一种跨膜糖蛋白,具有酪氨酸蛋白激酶活性,由细胞外N端与PDGF特异识别的结构域、单链顺序跨膜的中间疏水结构域和细胞内C端具有酪氨酸蛋白激酶活性的肽段结构域组成。当受体与其配体结合后促使两个受体分子形成二聚体,激活细胞内结构域酪氨酸残基自身磷酸化,或促使激活特殊靶蛋白的酪氨酸残基磷酸化,从而将信号传入细胞内,经级联式放大瀑布效应调控细胞的生命活动,包括靶细胞的分裂增殖[6]。
2 PDGF生物学效应和在肝纤维化中的作用
实验证明PDGF是一种重要的促有丝分裂因子,具有刺激特定细胞群分裂增殖的能力。肝受损时,大量分泌的PDGF刺激间质星形细胞增殖,转化为肌纤维样母细胞,并促使星形细胞迁移,聚集于炎症受损区[7]。而肌纤维样母细胞合成大量细胞外基质沉积于肝细胞胞间质,促进肝纤维化发生。
PDGF能够促进肌纤维母细胞产生胶原,尤其是I型及Ⅲ型胶原。Isbrucker[1]在体外培养的实验中证明,PDGF可促使未融合猪肝肌纤维样母细胞增殖,使已融合的细胞产生胶原,但对其细胞数量无影响。
PDGF还可通过上调组织金属蛋白酶抑制剂(TIMP-1)抑制胶原酶的作用,以减少细胞外基质的降解[4,8]。
3 PDGF与抗肝纤维化
3.1 PDGF的细胞内信号传导
抑制PDGF的产生和阻断其促纤维化的作用可望成为抗肝纤维化的途径,晚近通过对PDGF与其受体结合后细胞内信号传导途径的研究,发现阻断其信号传导能阻断PDGF的作用[9]。
PDGF与其受体结合后,可激活细胞内酪氨酸蛋白激酶(TPK),PDGF受体发生自动磷酸化,受体位点上的磷酸化酪因同源区2(Src-SH2)或酪氨酸磷酸区域进行结合[10]。结合于已激活的PDGF受体的分子包括有磷酯酰肌醇三激酶(PI 3-K),细胞外信号调节激酶(ERK)、磷酯酶C、酪氨酸激酶及Crb2、shc、shb等。经过一系列磷酸化反应,可活化核膜上丝氨酸(Ser)或苏氨酸(Thr)残基激酶,后作用于核内蛋白转录因子,如癌基因fos、jun、myc等编码的蛋白质,经直线传递信号使DNA的一些活性转录区参与基因表达的调控,或增加拓扑异构酶活力,改变细胞RNA含量,使细胞发生一系列表型改变,促细胞生长和分裂[7]。(如图1)。
图1 PDGF的细胞内信号传导途径
在PDGF发挥的作用中,PI 3-K及ERK途径是膜内信号传导的两个最重要途径。同时,PDGF引起细胞[Ca2+]浓度增加也促进了细胞增殖[6]。
3.2 细胞外信号调节激酶(ERK)及其抑制剂
业经证明ERK激活对激发细胞增殖是必不可少的。同时ERK途径与细胞外基质成分胶原的产生有密切关系[11]。ERK有两种构型,即ERK1(P44MAPK)和ERK2(P42MAPK)。人类肝星形细胞在ERK激活后继有c-fos激活,进一步作用于DNA转录因子,促细胞分裂[12]。故对ERK的作用进行干预抑制PDGF诱导的HSC增殖。若预先将HSC与升高细胞内cAMP水平的药物孵育,能明显降低PDGF诱导的ERK磷酸化、ERK活性、c-fos表达及促有丝分裂作用。己酮可可碱(Pentoxiffilly, PTF)是一种磷酸二酯酶抑制剂,可促细胞内cAMP浓度升高,抑制ERK传导途径和细胞增殖[13]。Preaux[14]实验证明,PTF可以减少PDGF诱导的细胞增殖,抑制Ⅰ、Ⅲ型胶原的合成,并使Ⅰ、Ⅲ型胶原mRNA减少,说明抑制胶原合成发生在转录前水平。
3.3 磷酯酰醇三激酶(PI 3-K)及其抑制剂
PI 3-K聚集激活PDGF受体,它由85KD的调节亚单位(2个SH-2区域)和110KD的催化亚单位构成。PDGF刺激PI 3-K与其受体结合,使85KD亚单位酪氨酸磷酸化后,激活蛋白激酶C和蛋白激酶B。这一信号传导引起细胞增殖及迁移,并聚集于炎症区[15]。PDGF-AA、AB及BB均可激活PI 3-K,以PDGF-BB作用最强,这与其促细胞分裂增殖能力有关。PDGF-BB对HSC的增殖和化学趋化性起作 用,而PDGF-A只促HSC增殖,无化学趋化作用,这可能与结合于不同受体,以及与受体后信号传导途径不同有关。渥曼青霉素(wortmannin)是一种真菌代谢产物,可结合PI 3-K,非竞争性抑制PI 3-K活性;但对受体的自动磷酸化和85KD蛋白与受体之间的联系无影响。渥曼青霉素还能部分抑制ERK活性,对它的深入研究,有可能成为一种新的抗纤维化药物[7]。
3.4 钙离子及其抑制剂
钙离子浓度的变化也与细胞增殖有关。实验证明PDGF明显升高细胞内[Ca2+][17]。其机理可能为PDGF与其受体结合后,酪氨酸激酶自动磷酸化,诱导磷酸肌醇(IP)循环,其循环产生IP3作用于细胞内皮网特异受体使其Ca2+释放,引起细胞内Ca2+水平迅速增加启动细胞内信号系统(I相)。IP3进一步磷酸化产生IP4又促使细胞外Ca2+内流 ,即细胞内Ca2+浓度的慢反应(Ⅱ相)[16]。证实PDGF诱导的Ca2+升高主要为细胞外Ca2+内流,与“T”型通道打开有关[17]。发现有机物氯化镍抑制“T”型通道,故有可能抑制PDGF所诱导的有丝分裂。因此,开发“T”通道特异抑制剂可阻断肝纤维化[9]。国内学者[18]选用细胞外钙离子拮抗剂维拉帕米(ver)和细胞内钙离子拮抗剂尼卡地平(nic),观察其对大鼠肝细胞胶原合成及升钙作用,结果显示ver可抑制PDGF促胶原合成和升钙效用,若ver与nic合用效果更佳。
3.5 细胞Na+-H+交换途径及其抑制
Di Sario[19]等人的研究表明:HSC激活伴随着Na+-H+交换的激活,细胞内pH增加。这种激活依赖于钙离子-钙调蛋白及蛋白激酶C途径。证明PDGF可增加Na+-H+交换活性,促进细胞分裂。Na+-H+交换抑制剂阿米洛利(amiloride)可抑制PDGF诱导的细胞增殖,故可能有抗纤维化作用。
3.6 酪氨酸蛋白激酶(TPK)及其抑制剂
PDGF与受体结合,激活受体细胞膜内部分的TPK,引起一系列信号传导及调节细胞增殖。因此阻断PDGF作用的关键之一是阻断TPK的活性。槲皮素(quercetin)是一种TPK抑制剂,属黄酮类,可以阻断细胞膜内酪氨酪残基的磷酸化使TPK失活,PDGF不能发挥作用[20]。故对TPK抑制剂的开发可望成为阻断纤维化的另一途径。
参考文献:
[1]Isbrucker RA,Oeferson Tc.Platelet-derived growth factor and pentoxifylline modulation of collagen synthesis in myofibroblasts.Toxicology and Applied[J].Pharmacology,1998,149(1)∶120
[2] Ikura Y,Morimoto H,Ogami M ,et al.Expression of platelet-derived growth factor and its receptor in livers of patients with chronic liver disease[J].J Castroentrology,1997,32(4)∶496
[3] Pinani M,Milani S, Herbst H ,et al.Expression of platelet-derived growth factor and its receptors in normal human liver and during active hepatic fibrosis[J].Am J Pathol,1996,148(3)∶785
[4] Burt AD,Cellular and molecular aspects of hepatic fibrosis[J].J Pathology,1993,170(2)∶105
[5] Wang L,Yamasaki G, Johnso R ,et al.Induction of β-platelet-derived growth factor receptor in rat hepatic lipocytes during cellular activation in vivo and in culture[J].J Clin Invest,1994,94(4)∶1563
[6] Friedman SL.Closing in on the signals of hepatic fibrosis[J].Grastroenterology,1997,112(4)∶1406
[7]Marra F,Gentilini A,Pinzani M,et al.Phosphatidylinositol 3-kinase is requried for plateled-derived growth factor′s actions on hepatic stellate cells[J].Gastroenterology,1997,112(4)∶1297
[8] Herbst H ,Schupan D,Milani S.Fibrogenesis and fibrolysis in ths liver[J].Verh Dtsch Ges Pathol,1995,79(1)∶15
[9] Pinzanj M,Marra F, Carlonj V.Signal transduction in hepatic stellate cells[J].Liver,1998,18(1)∶2
[10] Pawso T, Protein modules and signalling networks[J].Nature, 1995,373(4)∶573
[11] Davis BH,Chen A,Beno DWA.Raf and mitogen-activated protein kinase regulate stellate cell collagen gene expression[J].J Biol Chem,1996,271(3)∶11039
[12]Marra F,Ghosh choudhury G,Abboud HE.Interferon-gamma mediated activation of statl α regulates growth factor induced mitogenesis[J].J Clin Invest,1996,98(3)∶1218
[13] Pinznj M,Marra F,Gallgiuki A ,et al.Inhibiton by Pentoxifylline of extracelluar signal-regulated kinase activaton by plateletderived growth factor in hepatic stellate cells[J].Br J Pharmacol,1996,119(6)∶1117
[14] Preaux AM, Mallat A, Rosenbaum J,et al.Pentoxiffyline lnhibits growth and collagen synthesis of cultured human hepatic myofibrobloast-like cells[J].Hepatology,1997,26(2)∶315
[15] Valius M,Kazlaukas A.Phospholipase C-gamma and phosphatidylinostiol 3-kinase are the downstream mediators of the PDGF receptor′s mitogenic signal[J].Cell,1993,73(1)∶321
[16] Failli P,Ruocco C, Franco RD ,et al.The mitogenic effect of platelet-derived growth factor in human hepatic stellate cells requires calcium influx[J].Am J Physiol,1995,269(38)∶C1133
[17] Wang Z, Estacion M, Mordan LJ.Ca2+ Influx via-T type channels modulates PDGF-Induced replication of mouse fibroblasts[J].Am J Physiol,1993,265(39)∶C1239
[18] 姚迪,李定国,徐芹芳.血小板衍生生长因子和钙离子拮抗剂对大鼠肝细胞胶原合成及细胞内游离钙浓度的影响[J].胃肠病学和肝病学杂志,1998,7(2)∶132
[19] Di Sario A,Sregliati Baroni G, Bendia E, et al.Characterization of ion transport mechanisms regulating intracellular pH in hepatic stellate cells[J].Am J Physiol ,1997,273(36)∶G39
[20] Levitzki A, Gazit A.Tytosine kinase inhibition:an approach to drug development[J].Science, 1995,26T(5)∶1782
(收稿日期:1999-12-08 修回日期:2000-01-10)
您现在查看是摘要介绍页,详见PDF附件(180kb)。