贮脂细胞TGF-β1反义基因转移及对细胞外基质合成的抑制作用
作者:梁志清 何振平
单位:400038 重庆,第三军医大学西南医院肝胆外科
关键词:转化生长因子β;基因;反义;细胞外基质
中华医学杂志981121 【摘要】 目的 探讨TGF-β1在肝纤维化贮脂细胞激活和细胞外基质产生中的作用。以及对细胞外基质基因的表达和自身mRNA表达的作用。方法 将TGF-β1的基因序列反向插入逆转录病毒载体pLXSN,构建反义TGF-β1的逆转录病毒载体pLATSN,用PA317包装细胞包装,获得高滴度的逆转录病毒。将携有反义TGF-β1cDNA的逆转录病毒载体pLATSN导入人的贮脂细胞株LI90内,选择稳定转染的贮脂细胞株培养,应用PCR、RT-PCR检测反义基因的表达,用ELISA、免疫组织化学和原位杂交等方法检测TGF-β1和细胞外基质的产生。结果 贮脂细胞株LI90内有反义TGF-β1的表达,且其合成分泌TGF-β1和细胞外基质如FN、ColA1降低。结论 反义TGF-β1RNA可以抑制贮脂细胞的激活和内源性TGF-β1和细胞外基质的产生,为抗纤维化基因治疗提供理论基础。
, http://www.100md.com
TGF-β1 antisense gene transfer into ito cells and suppressed extracellular matrix production Liang Zhiqing, He Zhenping. Hepatobiliary Surgery Center, Southwestern Hospital, Third Military Medical University, Chongqing 400038
【Abstract】 Objective To investigate possible role of antisense TGF-β1 RNA in the regulation of TGF-β1 and ECM production in Ito cells. Method A human transforming growth factor-β1(TGF-β1) cDNA (1467 bp) was inserted in reverse orientation into the retroviral vector, constructed retroviral vector pLATSN of antisense RNA for TGF-β1. A high-titer, recombinant retroviral vector carried antisense RNA for TGF-β1 produced in PA317 packaging cells has been introduced into human Ito cells lines LI90. After selection with G418, resistant colonies were obtained. Results Stable integration of retrovirus in infectants was shown the presence of antisense RNA was detected by RT-PCR. The expression of TGF-β1 protein and the production of extracellular matrix such as FN, ColA1 were markedly decreased in the antisense TGF-β1 transfected cultured cells by ELISA, immunohistochemistry and in situ hybridization. Conclusion Antisense RNA of TGF-β1 can be successfully used to inhibit Ito cells activated, endogenous TGF-β1 mRNA and extracellular matrix produced, and may provide a basis for the development of anti-fibrosis gene therapy.
, http://www.100md.com
【Key words】 Transforming growth factor-beta Gene Antisense Extracellular matrix
(Natl Med J China, 1998, 78:850-852)
转化生长因子-β1(TGF-β1)在肝纤维化的发生、发展中起重要作用,除了激活Ito细胞合成细胞外基质,还能抑制肝细胞的再生。同时Ito细胞又能表达TGF-β1,通过自分泌和旁分泌作用,上调TGF-β1的表达,从而加速肝纤维化 的进程[1~3]。因此通过抑制TGF-β1的产生或阻断其生物活性来防治肝纤维化,是值得研究的领域[4]。我们根据TGF-β1的编码序列设计了其反义RNA的表达载体,并导入Ito细胞株LI90内,观察了其抑制TGF-β1表达及细胞外基质合成的效应,现将结果报道如下。
材料与方法
, http://www.100md.com
一、材料
克隆于重组逆转录病毒载体的多克隆位点的含人开放阅读框的反义TGF-β1cDNA质粒载体pLATSN,由本室构建。病毒包装细胞PA317由本校分子生物教研室刘昕博士惠赠,人Ito细胞株由日本东京大学医学院病理系的Murakami博士赠送。脂质体转染试剂盒购自Biolab,G418、Polybrene购自美国Sigma公司。TGF-β1抗体及标准品购自美国Promage公司,地高辛标记DNA及检测试剂盒由Boehringer Mannhem公司提供。
二、方法
1.重组病毒的包装及滴度测定:用脂质体法将pLATSN导入病毒包装细胞PA317中,经G418筛选获得抗性克隆,用NIH3T3细胞测定重组病毒的滴度,挑选高滴度的PA317细胞克隆收集其上清,转导人贮脂细胞LI90。随后在长至70%瓶底的LI90细胞中加入含8 μg/ml Polybrene的病毒上清液0.5 ml,均匀覆盖瓶底,每隔30分钟轻摇1次,至2小时加入3 ml的新鲜培养液,24小时后1∶2传代并加入含有250 μg/mlG418的培养液,再过48小时改用420 μg/ml G418的培养液,筛选2周获得阳性克隆。
, http://www.100md.com
2.外源基因在贮脂细胞LI90中整合的鉴定:(1) 细胞DNA及RNA的提取按试剂盒说明书进行,PCR引物的设计,TGF-β1 cDNA上游引物P1:5′-GGG ACT ATC CAC CTG CAA GA-3′;下游引物P2:5′-AGG ACC TTG CTG TAC TGC GT-3′。(2) RT-PCR检测反义TGF-β1RNA表达:反转录PCR(RT-PCR)均按试剂盒说明书操作,PCR反应条件如下(PE公司9600型PCR仪):95 ℃变性3分钟,下述条件循环30次:95 ℃1分钟,60 ℃1分钟,72 ℃2分钟。
3.TGF-β1生物活性的测定:基因转导的贮脂细胞LI90经G418筛选获得的抗性克隆,随后扩大培养,加入无血清培养液过夜48小时,收集上清,用ELISA法测定TGF-β1的含量,以标准品的比活性计算TGF-β1的活性。
4.反义TGF-β1RNA对贮脂细胞细胞外基质合成的影响:采用原位杂交和免疫组织化学检测FN、和ColA1的变化,按常规方法进行。
, 百拇医药
结果
一、包装细胞重组病毒滴度测定
经Lipofect AMINE介导,将pLATSN质粒和空载体质粒pLXSN转染PA317细胞,以G418筛选获得抗性克隆,分别命名为PALATSN和PALXSN。随机挑选6个克隆,收集其上清,感染NIH 3T3细胞测定共病毒滴度,空载体病毒滴度为6.85×104 CFU/ml,重组载体病毒滴度为4.34×104CFU/ml。
二、反义TGF-β1基因在贮脂细胞中的稳定表达
应用重组病毒PALATSN上清感染贮脂细胞LI90,挑取G418抗性克隆,提取细胞RNA,采用RT-PCR扩增反义RNA:选用下游引物(3′端引物),即P2,做为反转录引物,先将细胞内的反义RNA反转录成cDNA,用P1和P2扩增反转录检测细胞内是否有反义RNA,可见PALATSN感染细胞中可扩增出939 bp的反义RNA,而未感染细胞中则未扩增出该片段(图1)。
, http://www.100md.com
M:λDNA/HindⅢ,1.LI90, 2,3.LIAT
图1 转基因细胞RT-PCR扩增发现有939 bp带,未转基因细胞缺乏此带
三、培养细胞上清TGF-β1活性
对TGF-β1反义基因转染后G418抗性克隆细胞扩大培养后发现,细胞的生长能力明显减低甚至部分细胞死亡。测定培养细胞上清TGF-β1活性,发现重组病毒载体转染的6个G418抗性贮脂细胞克隆1、2、3、4、5和6,其分泌的TGF-β1分别为33.5、58、71.6、23.1、69.1和52.1(每48小时ng/ml),明显低于空载体的3个阳性克隆1、2和3分泌的TGF-β1量即124.3、137.3和98.7(每48小时ng/ml),及未转染转染细胞分泌的TGF-β1量为142.7(每48小时ng/ml)。
四、反义TGF-β1 RNA对贮脂细胞细胞外基质合成的影响。
, http://www.100md.com
原位杂交及免疫组化显示,转染反义TGF-β1的贮脂细胞表达的FN、ColA1阳性细胞数及阳性程度明显低于未转染及空载体转染的贮脂细胞(图2~4)。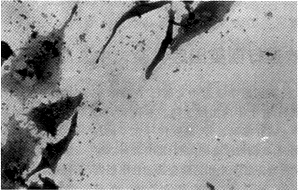
图2 未转基因细胞ColA1免疫组化染色为强阳性 SP×200
图3 转基因细胞ColA1免疫组化染色为弱阳性 SP×200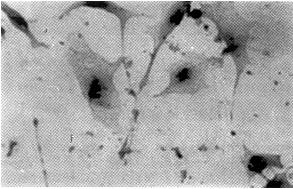
图4 未转基因细胞TGF-β1原位杂交为强阳性 SP×200
讨论
, 百拇医药 由于TGF-β1在肝纤维化中的重要作用,因此,如能抑制或阻断TGF-β1的产生和活性,有可能阻止肝纤维化的进程,从而达到防治肝纤维化的目的。就现有的方法多限于下列两个方面:一是利用抗TGF-β1抗体中和TGF-β1的活性,这方面的研究已初步取得满意的结果[5];二是采用间接的手段调节TGF-β1的活性,如中医中药、TGF-β1拮抗剂等[6],但效果不确切。
为了能从基因水平抑制、阻断TGF-β1的产生,我们应用反义技术,将TGF-β1基因片段反向构建于逆转录病毒载体,用脂质体介导转入包装细胞PA317,再将包装细胞产生的重组病毒感染贮脂细胞,使反义TGF-β1 RNA整合入贮脂细胞,使其在基因转录、翻译水平阻止贮脂细胞TGF-β1的表达。通过RT-PCR发现反义TGF-β1 RNA已整合入贮脂细胞基因组,用ELISA法检测发现转基因阳性克隆细胞TGF-β1活性明显低于正常细胞,表明反义TGF-β1 RNA能使贮脂细胞合成分泌TGF-β1减少或消失。采用原位杂交及免疫组化发现,转基因贮脂细胞表达FN、ColA1的阳性细胞及阳性程度明显低于未转染及空载体转染细胞。因此,我们认为反义TGF-β1 RNA不仅可以阻止自身TGF-β1的合成、分泌以减弱或消除对贮脂细胞的自分泌调节作用,还能阻止或减少Ito细胞胶原等细胞外基质的合成[7],以减缓或阻止肝纤维化的进程。
, 百拇医药
本课题为国家自然科学基金资助项目
参考文献
1 Czaja MJ, Weiner FR, Flanders KC, et al. In vitro and in vivo association of transforming growth factor-beta with hepativ fibrosis. J Cell Biol, 1989, 108:2477-2482.
2 Annoni G, Weiner FR, Zern MA. Increased transforming growth factor-beta (TGF-beta) gene expression in human liver disease. J Hepatol, 1992, 14:259-264.
3 Casini A, Pinzani M, Milani S, et al. Regulation of extracellular matrix synthesis by transforming growth factor-beta1 in human fat-storing cells. Gastroenterology, 1993, 105:245-253.
, 百拇医药
4 Trotter JF, Brenner DA. Current and prospetive therpies for hepatic fibrosis. Compre Ther, 1995, 21:303-307.
5 Strobel D, Muller P, Reiser K, et al. Regulation of collagen expression in vivo in experimental liver fibrosis by a neutralizing TGF-beta antibody. Hepatology, 1994, 20:293-302.
6 Trotter JF, Brenner DA, Gentry L, et al. Latency associated peptide (LAP), an inhibitor of transforming growth factor-beta, as a therapy for experimental hepatic fibrosis. Hepatology, 1994, 20:227-238.
7 Akagi Y, Isaka Y, Kaneko T, et al. Inhibition of TGF-beta1 expression by antisense oligonucleotides suppressed extracellular matrix accumulation in experimental glomeruphritis. Kidney Int, 1996, 50:148-155.
(收稿:1997-10-22 修回:1998-08-06), http://www.100md.com
单位:400038 重庆,第三军医大学西南医院肝胆外科
关键词:转化生长因子β;基因;反义;细胞外基质
中华医学杂志981121 【摘要】 目的 探讨TGF-β1在肝纤维化贮脂细胞激活和细胞外基质产生中的作用。以及对细胞外基质基因的表达和自身mRNA表达的作用。方法 将TGF-β1的基因序列反向插入逆转录病毒载体pLXSN,构建反义TGF-β1的逆转录病毒载体pLATSN,用PA317包装细胞包装,获得高滴度的逆转录病毒。将携有反义TGF-β1cDNA的逆转录病毒载体pLATSN导入人的贮脂细胞株LI90内,选择稳定转染的贮脂细胞株培养,应用PCR、RT-PCR检测反义基因的表达,用ELISA、免疫组织化学和原位杂交等方法检测TGF-β1和细胞外基质的产生。结果 贮脂细胞株LI90内有反义TGF-β1的表达,且其合成分泌TGF-β1和细胞外基质如FN、ColA1降低。结论 反义TGF-β1RNA可以抑制贮脂细胞的激活和内源性TGF-β1和细胞外基质的产生,为抗纤维化基因治疗提供理论基础。
, http://www.100md.com
TGF-β1 antisense gene transfer into ito cells and suppressed extracellular matrix production Liang Zhiqing, He Zhenping. Hepatobiliary Surgery Center, Southwestern Hospital, Third Military Medical University, Chongqing 400038
【Abstract】 Objective To investigate possible role of antisense TGF-β1 RNA in the regulation of TGF-β1 and ECM production in Ito cells. Method A human transforming growth factor-β1(TGF-β1) cDNA (1467 bp) was inserted in reverse orientation into the retroviral vector, constructed retroviral vector pLATSN of antisense RNA for TGF-β1. A high-titer, recombinant retroviral vector carried antisense RNA for TGF-β1 produced in PA317 packaging cells has been introduced into human Ito cells lines LI90. After selection with G418, resistant colonies were obtained. Results Stable integration of retrovirus in infectants was shown the presence of antisense RNA was detected by RT-PCR. The expression of TGF-β1 protein and the production of extracellular matrix such as FN, ColA1 were markedly decreased in the antisense TGF-β1 transfected cultured cells by ELISA, immunohistochemistry and in situ hybridization. Conclusion Antisense RNA of TGF-β1 can be successfully used to inhibit Ito cells activated, endogenous TGF-β1 mRNA and extracellular matrix produced, and may provide a basis for the development of anti-fibrosis gene therapy.
, http://www.100md.com
【Key words】 Transforming growth factor-beta Gene Antisense Extracellular matrix
(Natl Med J China, 1998, 78:850-852)
转化生长因子-β1(TGF-β1)在肝纤维化的发生、发展中起重要作用,除了激活Ito细胞合成细胞外基质,还能抑制肝细胞的再生。同时Ito细胞又能表达TGF-β1,通过自分泌和旁分泌作用,上调TGF-β1的表达,从而加速肝纤维化 的进程[1~3]。因此通过抑制TGF-β1的产生或阻断其生物活性来防治肝纤维化,是值得研究的领域[4]。我们根据TGF-β1的编码序列设计了其反义RNA的表达载体,并导入Ito细胞株LI90内,观察了其抑制TGF-β1表达及细胞外基质合成的效应,现将结果报道如下。
材料与方法
, http://www.100md.com
一、材料
克隆于重组逆转录病毒载体的多克隆位点的含人开放阅读框的反义TGF-β1cDNA质粒载体pLATSN,由本室构建。病毒包装细胞PA317由本校分子生物教研室刘昕博士惠赠,人Ito细胞株由日本东京大学医学院病理系的Murakami博士赠送。脂质体转染试剂盒购自Biolab,G418、Polybrene购自美国Sigma公司。TGF-β1抗体及标准品购自美国Promage公司,地高辛标记DNA及检测试剂盒由Boehringer Mannhem公司提供。
二、方法
1.重组病毒的包装及滴度测定:用脂质体法将pLATSN导入病毒包装细胞PA317中,经G418筛选获得抗性克隆,用NIH3T3细胞测定重组病毒的滴度,挑选高滴度的PA317细胞克隆收集其上清,转导人贮脂细胞LI90。随后在长至70%瓶底的LI90细胞中加入含8 μg/ml Polybrene的病毒上清液0.5 ml,均匀覆盖瓶底,每隔30分钟轻摇1次,至2小时加入3 ml的新鲜培养液,24小时后1∶2传代并加入含有250 μg/mlG418的培养液,再过48小时改用420 μg/ml G418的培养液,筛选2周获得阳性克隆。
, http://www.100md.com
2.外源基因在贮脂细胞LI90中整合的鉴定:(1) 细胞DNA及RNA的提取按试剂盒说明书进行,PCR引物的设计,TGF-β1 cDNA上游引物P1:5′-GGG ACT ATC CAC CTG CAA GA-3′;下游引物P2:5′-AGG ACC TTG CTG TAC TGC GT-3′。(2) RT-PCR检测反义TGF-β1RNA表达:反转录PCR(RT-PCR)均按试剂盒说明书操作,PCR反应条件如下(PE公司9600型PCR仪):95 ℃变性3分钟,下述条件循环30次:95 ℃1分钟,60 ℃1分钟,72 ℃2分钟。
3.TGF-β1生物活性的测定:基因转导的贮脂细胞LI90经G418筛选获得的抗性克隆,随后扩大培养,加入无血清培养液过夜48小时,收集上清,用ELISA法测定TGF-β1的含量,以标准品的比活性计算TGF-β1的活性。
4.反义TGF-β1RNA对贮脂细胞细胞外基质合成的影响:采用原位杂交和免疫组织化学检测FN、和ColA1的变化,按常规方法进行。
, 百拇医药
结果
一、包装细胞重组病毒滴度测定
经Lipofect AMINE介导,将pLATSN质粒和空载体质粒pLXSN转染PA317细胞,以G418筛选获得抗性克隆,分别命名为PALATSN和PALXSN。随机挑选6个克隆,收集其上清,感染NIH 3T3细胞测定共病毒滴度,空载体病毒滴度为6.85×104 CFU/ml,重组载体病毒滴度为4.34×104CFU/ml。
二、反义TGF-β1基因在贮脂细胞中的稳定表达
应用重组病毒PALATSN上清感染贮脂细胞LI90,挑取G418抗性克隆,提取细胞RNA,采用RT-PCR扩增反义RNA:选用下游引物(3′端引物),即P2,做为反转录引物,先将细胞内的反义RNA反转录成cDNA,用P1和P2扩增反转录检测细胞内是否有反义RNA,可见PALATSN感染细胞中可扩增出939 bp的反义RNA,而未感染细胞中则未扩增出该片段(图1)。
, http://www.100md.com
M:λDNA/HindⅢ,1.LI90, 2,3.LIAT
图1 转基因细胞RT-PCR扩增发现有939 bp带,未转基因细胞缺乏此带
三、培养细胞上清TGF-β1活性
对TGF-β1反义基因转染后G418抗性克隆细胞扩大培养后发现,细胞的生长能力明显减低甚至部分细胞死亡。测定培养细胞上清TGF-β1活性,发现重组病毒载体转染的6个G418抗性贮脂细胞克隆1、2、3、4、5和6,其分泌的TGF-β1分别为33.5、58、71.6、23.1、69.1和52.1(每48小时ng/ml),明显低于空载体的3个阳性克隆1、2和3分泌的TGF-β1量即124.3、137.3和98.7(每48小时ng/ml),及未转染转染细胞分泌的TGF-β1量为142.7(每48小时ng/ml)。
四、反义TGF-β1 RNA对贮脂细胞细胞外基质合成的影响。
, http://www.100md.com
原位杂交及免疫组化显示,转染反义TGF-β1的贮脂细胞表达的FN、ColA1阳性细胞数及阳性程度明显低于未转染及空载体转染的贮脂细胞(图2~4)。
图2 未转基因细胞ColA1免疫组化染色为强阳性 SP×200
图3 转基因细胞ColA1免疫组化染色为弱阳性 SP×200
图4 未转基因细胞TGF-β1原位杂交为强阳性 SP×200
讨论
, 百拇医药 由于TGF-β1在肝纤维化中的重要作用,因此,如能抑制或阻断TGF-β1的产生和活性,有可能阻止肝纤维化的进程,从而达到防治肝纤维化的目的。就现有的方法多限于下列两个方面:一是利用抗TGF-β1抗体中和TGF-β1的活性,这方面的研究已初步取得满意的结果[5];二是采用间接的手段调节TGF-β1的活性,如中医中药、TGF-β1拮抗剂等[6],但效果不确切。
为了能从基因水平抑制、阻断TGF-β1的产生,我们应用反义技术,将TGF-β1基因片段反向构建于逆转录病毒载体,用脂质体介导转入包装细胞PA317,再将包装细胞产生的重组病毒感染贮脂细胞,使反义TGF-β1 RNA整合入贮脂细胞,使其在基因转录、翻译水平阻止贮脂细胞TGF-β1的表达。通过RT-PCR发现反义TGF-β1 RNA已整合入贮脂细胞基因组,用ELISA法检测发现转基因阳性克隆细胞TGF-β1活性明显低于正常细胞,表明反义TGF-β1 RNA能使贮脂细胞合成分泌TGF-β1减少或消失。采用原位杂交及免疫组化发现,转基因贮脂细胞表达FN、ColA1的阳性细胞及阳性程度明显低于未转染及空载体转染细胞。因此,我们认为反义TGF-β1 RNA不仅可以阻止自身TGF-β1的合成、分泌以减弱或消除对贮脂细胞的自分泌调节作用,还能阻止或减少Ito细胞胶原等细胞外基质的合成[7],以减缓或阻止肝纤维化的进程。
, 百拇医药
本课题为国家自然科学基金资助项目
参考文献
1 Czaja MJ, Weiner FR, Flanders KC, et al. In vitro and in vivo association of transforming growth factor-beta with hepativ fibrosis. J Cell Biol, 1989, 108:2477-2482.
2 Annoni G, Weiner FR, Zern MA. Increased transforming growth factor-beta (TGF-beta) gene expression in human liver disease. J Hepatol, 1992, 14:259-264.
3 Casini A, Pinzani M, Milani S, et al. Regulation of extracellular matrix synthesis by transforming growth factor-beta1 in human fat-storing cells. Gastroenterology, 1993, 105:245-253.
, 百拇医药
4 Trotter JF, Brenner DA. Current and prospetive therpies for hepatic fibrosis. Compre Ther, 1995, 21:303-307.
5 Strobel D, Muller P, Reiser K, et al. Regulation of collagen expression in vivo in experimental liver fibrosis by a neutralizing TGF-beta antibody. Hepatology, 1994, 20:293-302.
6 Trotter JF, Brenner DA, Gentry L, et al. Latency associated peptide (LAP), an inhibitor of transforming growth factor-beta, as a therapy for experimental hepatic fibrosis. Hepatology, 1994, 20:227-238.
7 Akagi Y, Isaka Y, Kaneko T, et al. Inhibition of TGF-beta1 expression by antisense oligonucleotides suppressed extracellular matrix accumulation in experimental glomeruphritis. Kidney Int, 1996, 50:148-155.
(收稿:1997-10-22 修回:1998-08-06), http://www.100md.com