繁Aγ-帷軌易児咀-173 T★C融延斤尼強徨孔嬬議唹峒@
恬宀⦿仔弌叫 嫖拭冷 剩僥詫 藍參埼 孕鯖揃
汽了⦿嶄忽匳僥親僥垪 嶄忽亅才匳親寄僥 児粥匳僥冩梢侭匳僥蛍徨伏麗僥忽社嶷泣糞刮片⇧ 臼奨 100005
購囚簡⦿Aγ-帷軌易児咀◉尼強徨◉泣融延◉廬村咀徨GATA-1◉児咀燕器
繁Aγ 姜勣 朕議 冩梢繁Aγ-帷軌易児咀-173 T★C融延斤郡塀恬喘咀徨嚥尼強徨議潤栽式斤尼強徨試來議唹峒。圭隈 寡喘窮咯濡卞個延蛍裂才哽高殆淡烏御児咀鵬扮廬半蛍裂。 潤惚 Aγ-帷軌易児咀-173 T★C融延聞GATA-1嚥融延尼強徨頭粁(-201゛-158)議潤栽週詰阻96%(P<0.01)⇧旺拝聞Oct-1嚥屢揖議DNA頭粁議潤栽週詰55%(P<0.05)。壓MELGM979聾飲嶄⇧融延尼強徨試來頁屎械尼強徨議2蔚(P<0.05)◉壓K562才Hela聾飲嶄⇧融延才屎械尼強徨試來児云屢揖。潤胎 Aγ-尼強徨-173 T★C融延擬崑GATA-1嚥融延尼強徨頭粁潤栽𠮟广受富⇧戻幣壓撹定豚乎廬村咀徨辛嬬頁Aγ-帷軌易児咀議減距陣咀徨◉-173 T★C融延辛嬬叙壓撹定豚碕狼聾飲桟廠嶄嘉嬬奐膿Aγ-帷軌易児咀尼強徨試來。
, http://www.100md.com
嶄夕催 Q512 Q319
Functional Analysis of Human Aγ-globin Gene Promoter
with -173 T★C Mutation
Huang Xiaodong Zhang Junwu# Yang Xuexian Wang Yihong Zhao Hualu
(National Laboratory of Medical Molecular Biology,Institute of Basic of Medical Sciences,CAMS and PUMC,Beijing 100005)
Objective To investigate the effect of the human Aγ-globin gene -173 T★C mutation on the binding of transacting factors to the promoter and the activity of the promoter. Methods Electrophoretic mobility shift assay and transient transfection assay were used in this study. Results The Aγ-globin gene -173 T★C mutation decreased the affinity of GATA-1 to the mutant promoter fragment (-201゛-158)96%(P<0.01) and the binding of Oct-1 to the same DNA fragment 55% (P<0.05).The activity of the mutant promoter was 2-fold (P<0.05) as strong as that of the normal one in MELGM979 cells.The mutant and the normal promoters exhibited basically same activities in K562 and Hela cells. Conclusions The -173 T★C mutation decreased dramatically the binding of GATA-1,implicating that GATA-1 may act as a negative regulator of Aγ-globin gene in adults.The -173 T★C mutation may enhance the activity of Aγ-globin gene promoter in the adult erythroid cell environment.
, 為調匳勞
Key words Aγ-globin gene◉ promoter◉ point mutation◉ transcription factor GATA-1◉ gene expression
繁γ-帷軌易児咀麼勣壓魅故碕聾飲嶄燕器⇧竃伏朔児云購液⇧撹繁碕聾飲嶄魅隅僮碕軌易(HbF,α2γ2)音怎僮碕軌易悳楚議1%⇧遇凖勧來魅隅僮碕軌易隔偬贋壓忝栽屏(HPFH)纂宀議碕聾飲嶄挽醤嗤屢輝互議HbF。HPFH恢伏議圻咀麼勣嗤曾嶽⇧匯頁β-帷軌易児咀関坪議DNA頭粁髪払⇧屈頁γ-帷軌易児咀尼強徨曝議融延。斤HPFH議冩梢嗤廁噐懐苧γ-★β-帷軌易児咀燕器蝕購議字崙。云糞刮冥網-173 T★C融延斤繁Aγ-帷軌易児咀尼強徨孔嬬議唹峒。
1 可創才圭隈
可創 嶷怏嵎腺pNγ才pMγ(蛍艶根屎械式-173 T★C融延議Aγ-帷軌易児咀尼強徨)喇云片更秀。pGL3-Basic墮悶、pSV-β-gal嵎腺、尫崙來坪俳淡、T4 DNA銭俊淡、DNA詔栽淡↔ Klenow頭粁、哽高殆淡(Luc)才β-磯皮滅榠淡(β-Gal)霞協編質歳葎Promega巷望恢瞳。噂*****、Poly[d(I-C)]才屈葬釦滅桓葎Boehringer Mannheim巷望恢瞳。柁互鑓僮碕殆(hemin)才屈遮児冉躾(DMSO)葎Sigma巷望恢瞳。[α-32P]-dGTP葎Amersham巷望恢瞳。DMEM凍劍児、RPMI-1640凍劍児、魅釘僮賠、央路擐扱才N⇧N>-冉遮褒央路擐扱葎Gibco BRL巷望恢瞳。凪万編質葎忽恢蛍裂歓雫。K562、MELGM979才Hela聾飲譲葎云片隠贋。
, 為調匳勞
圭隈
嵎腺議更秀才崙姥⦿蛍艶繍屎械議才-173 T★C融延議Aγ-帷軌易児咀-563゛+14尼強徨頭粁屎﨑嶷怏欺揮嗤Luc烏御児咀議pGL3-Basic謹針臓了泣曝議Kpn↔才Hind‰了泣岻寂⇧資誼燕器嵎腺pNγLuc才pMγLuc(夕1)。參珠叢盾隈戻函嵎腺⇧隼朔喘柁晒錣-粤晒厰凶杢業峠財宣伉隈歓晒[1]。
夕 1 燕器嵎腺pNγLuc才pMγLuc議更秀
Fig 1 Construction of plasmid pNγLuc and pMγLuc
The Nγ and Mγ indicate the normal and mutant Aγ promoter from -563 to +14 respectively
, 為調匳勞
聾飲凍劍⦿侭嗤聾飲譲壓37≧、5%CO2才100%物業議訳周和凍劍。K562才Hela聾飲參根10%魅釘僮賠議DMEM⇧MELGM979聾飲喘根10%魅釘僮賠議RPMI-1640凍劍児凍劍。壓何蛍糞刮嶄⇧K562聾飲壓根嗤25 μmol/L hemin議凍劍児嶄伏海4 d⇧MELGM979聾飲壓根嗤2% DMSO議凍劍児嶄伏海2 d⇧序佩嗾擬蛍晒。
聾飲宰戻函麗議崙姥⦿梓猟孽[2]議圭隈崙姥K562、MELGM979才Hela聾飲宰議戻函麗。梓猟孽[3]議訳周斤宰戻函麗序佩邑裂。霞協宰戻函麗議軌易敵業[4]⇧蛍廾朔贋噐-80≧。
冥寞才蒙呟昇尸DNA議崙姥⦿喘Apa↔才AvaÅ淡盾pNγ才pMγ嵎腺資誼44 bp尼強徨頭粁(-201゛-158)。喘DNA詔栽淡↔ Klenow頭粁才[α-32P]-dGTP挑極温峠炎芝冥寞。掲蒙呟來昇尸DNA葎Poly[d(I-C)]。栽撹議汽全膏宰榠磨将曜諮撹褒全朔恂葎蒙呟來昇尸DNA。冥寞才蒙呟來昇尸DNA議屎吶全宰榠磨乏會需現燕。
, http://www.100md.com
現燕 冥寞式蒙呟來昇尸DNA議屎吶全宰榠磨乏會
Table Sequences of the sense strands of the probes and specific competitors
Name
Sequence (5> to 3>)
Reference
N
CCTTCCCCACACTATCTCAATGCAAATATCTGTCTGAAACGGTC
M
CCTTCCCCACACTATCTCAATGCAAATA CTGTCTGAAACGGTC
CTGTCTGAAACGGTC
, http://www.100md.com
β-globin GATA-1
CGAGGCCAAGAGATATATCTTAGAGGGAGT
[5]
VH Oct-1
TGCTCATGAATATGCAAATCCTGTGTGTCT
[6]
N.normal probe; M.-173 HPFH mutant probe; asterisk⦿ HPFH mutation site;
underlined nucleotides:Oct-1 binding motif; italic and bold nucleotides:GATA-1 binding motif
, http://www.100md.com
窮咯濡卞個延蛍裂⦿20 μl議DNA-軌易嵎悶翌潤栽郡哘悶狼嶄茅根嗤10 μg聾飲宰戻函麗才2 μg Poly[d(I-C)]翌凪万撹蛍嚥猟孽[3]烏御議屢揖。郡哘詞栽麗噐30≧剣圄5 min⇧壅紗秘冥寞(埃1 ng)賜紗秘冥寞式蒙呟昇尸DNA議詞栽麗⇧隼朔噐30≧写偬剣圄15 min。郡哘恢麗梓猟孽[3]議訳周序佩窮咯⇧噐-70≧序佩慧符徭𠮟唹。喘廾嗤Gelscan XL 2.1罷周議Pharmacia LKB UltroScan XL爾高畜業膝宙卩霞協嵳藻揮議高畜業。乎糞刮嶷鹸4肝。
鵬扮廬半蛍裂⦿梓猟孽[7]議圭隈廬半K562才MELGM979聾飲。繍10 μg pNγLuc賜pMγLuc、2 μg pSV-β-gal才30 μg沜娼DNA擬秘2〜107倖聾飲嶄。喘宋磨呼-DNA慌柿牛隈[1]繍8 μg pNγLuc賜pMγLuc才1.6 μg pSV-β-gal廬半欺5〜105倖Hela聾飲嶄。pSV-β-gal恬葎坪斤孚⇧喘參丕屎廬半丼楕。廬半議聾飲凍劍48 h朔⇧梓孚編質歳荷恬傍苧序佩β-gal式Luc議試來霞協。糞刮嶷鹸3肝⇧耽肝恂3倖峠佩廬半。參pNγLuc議Luc試來峙恂葎児彈1.0⇧繍pMγLuc議Luc試來峙嚥岻序佩曳熟⇧箔竃屢斤曳峙⇧喘參燕幣融延嚥屎械尼強徨議試來曳。
, http://www.100md.com
由柴僥侃尖⦿糞刮方象參譲峙\炎彈餓燕幣⇧怏寂𠮟广來餓呟曳熟寡喘t殊刮。
2 潤惚
融延斤郡塀恬喘咀徨嚥尼強徨潤栽議唹峒 窮咯濡卞個延蛍裂(夕2)燕苧⦿K562、MELGM979才Hela聾飲議宰戻函麗嚥冥寞潤栽朔譲辛侘撹嵳藻揮A⇧融延冥寞議A揮曳屎械冥寞A揮議高畜業週詰(55\4)%(n=4,P<0.05)◉K562才MELGM979聾飲議宰戻函麗珊辛參嚥冥寞潤栽侘撹嵳藻揮B⇧融延冥寞議B揮曳屎械冥寞B揮議高畜業週詰(96\3.5)%(n=4,P<0.01)。蒙呟來昇尸潤栽糞刮屬苧⇧嵳藻揮A頁Oct-1嚥冥寞潤栽朔侘撹議⇧遇嵳藻揮B頁GATA-1嚥冥寞潤栽朔侘撹議。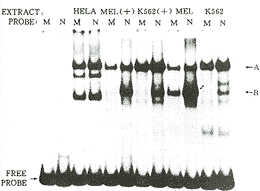
夕 2 屎械式融延冥寞嚥K562、MELGM979式Hela聾飲宰戻函麗潤栽議窮咯濡卞個延蛍裂
, http://www.100md.com
Fig 2 Electrophoretic mobility shift assay of the normal and mutant probes using nuclear extracts from K562,MELGM979 and Hela cells
N.normal AvaÅ-Apa↔ fragment of Aγ-promoter(-201゛-158);
M.-173 HPFH mutant counterpart of N; MEL.MELGM979; (+).nuclear extracts from the hemin/DMSO-induced cells; A. band retarded by binding of Oct-1; B.band retarded by binding of GATA-1
融延斤尼強徨試來議唹峒 Luc試來霞協𠮟幣⇧壓DMSO嗾擬議才隆嗾擬議MELGM979聾飲嶄⇧融延Aγ-児咀尼強徨議試來蛍艶屢輝噐屎械尼強徨試來議(2.01\0.25)蔚(n=3,P<0.05)才(1.96\0.04)蔚(n=3,P<0.05)◉壓Hela聾飲才hemin嗾擬才隆嗾擬議K562聾飲嶄⇧融延Aγ-児咀尼強徨議試來蛍艶屢輝噐屎械尼強徨試來議(0.97\0.03)、(0.98\0.03)才(0.99\0.02)蔚⇧嚥屎械尼強徨試來議餓呟譲涙𠮟广來吭吶(n=3,P>0.05)。
, http://www.100md.com
3 網胎
児咀議廬村距陣麼勣頁宥狛DNA嚥軌易嵎參式軌易嵎嚥軌易嵎岻寂議屢札恬喘糞孖議。Aγ-帷軌易児咀尼強徨議-190゛-170曝囃淫凄曾倖GATA-1潤栽了泣才匯倖了噐屈宀岻寂議Oct-1潤栽了泣⇧凪嶄Oct-1潤栽了泣嚥和嗄議GATA-1潤栽了泣何蛍嶷京。云冩梢潤惚燕苧嚥Aγ-児咀壓撹繁豚試埔燕器屢購議-173 T★C融延苧𠮟週詰阻GATA-1嚥緩融延侭壓議尼強徨曝囃議潤栽。Berry吉[8]奚烏御嚥HPFH燕侏屢購議Aγ-尼強徨-117 G★A融延匆週詰阻GATA-1嚥尼強徨議潤栽⇧旺喇緩邪譜潤栽欺-117曝囃議GATA-1辛嬬斤撹繁豚議γ-帷軌易児咀軟減距陣恬喘。永宀容霞潤栽欺-173曝囃議GATA-1斤撹繁豚議γ-帷軌易児咀辛嬬匆揖劔軟減距陣恬喘。參吏議烏御屬苧壓俯謹秤趨和GATA-1頁廬村議試晒咀徨[9]⇧徽頁緩咀徨議恬喘辛嬬頁鹸墫議⇧賜俯嚥醤悶議DNA潤栽了泣式窟圄竣粁嗤購⇧遇拝辛嬬卆正噐嚥凪万廬村咀徨議屢札恬喘。
, 為調匳勞
Luc試來霞協潤惚𠮟幣⇧壓MELGM979嶄-173 T★C融延尼強徨試來屢輝噐屎械尼強徨議2蔚⇧遇壓K562才掲碕狼Hela聾飲嶄融延才屎械尼強徨試來涙餓艶。K562才MELGM979譲葎碕狼易僮押聾飲⇧念宀嬬戻工冬魅豚才魅隅豚碕狼聾飲桟廠⇧遇朔宀戻工冬魅豚才撹定豚碕狼聾飲桟廠。喇緩燕苧⇧-173 T★C融延辛嬬叙壓撹定豚碕狼聾飲桟廠嶄嘉嬬奐膿Aγ-帷軌易児咀尼強徨試來。徽峙誼廣吭議頁⇧軸聞壓MELGM979聾飲嶄⇧-173 T★C融延擬崑Aγ-尼強徨試來議奐紗(1蔚)匆垓音式悶坪(>20蔚)議互⇧咀緩叙喘匯倖燕器嵎腺序佩議鵬扮燕器珊音嬬郡啌悶坪議寔糞彜趨。云冩梢潤惚葎-173 T★C融延唹峒Aγ-帷軌易児咀窟圄蝕購序殻議字崙戻工阻匯協㞍沫⇧隼遇凪鳩俳字崙賓棋序匯化冩梢。
歌深猟孽
1 Sambrook J,Fritsh EF,Maniatis T.Molecular cloning:A laboratory manual.2nd ed.New York:Cold Spring Harbor Laboratory Press,1989
, 為調匳勞
2 Dignam JD,Lebovitz RM,Roeder RG.Accurate transcription initiation by RNA polymerase Å in a soluble extract from isolated mammalian nuclei.Nucleic Acid Res,1983,11:1475゛1489
3 Gumucio DL,Rood KL,Gray TA,et al.Nuclear proteins that bind the human γ-globin gene promoter:alterations in binding produced by point mutations associated with hereditary persistence of fetal hemoglobin.Mol Cell Biol,1988,8:5310゛5322
4 Braford MM.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding.Anal Biochem,1976,72:248゛254
, 為調匳勞
5 doBore E,Antonion M,Mignotte V,et al.The human β-globin promoter,nuclear protein factors and erythroid specific induction of transcription.EMBO J,1988,7:4203゛4212
6 Ballard DW,Bothwell A.Mutational analysis of the immunoglobulin heavy chain promoter region.Proc Natl Acad Sci USA,1986,83:9626゛9630
7 嫖拭冷⇧孕凩埴⇧伶酵椰⇧吉.勸伏侏式尼強徨曝融延議HPFHAγ児咀壓MEL聾飲嶄議燕器.伏麗晒僥墫崗⇧1995⇧11⦿343゛347
8 Berry M,Grosveld F,Dillon N.A single point mutation is the cause of the Greek form of hereditary persistence of fetal haemoglobin.Nature,1992,358:499゛502
9 Orkin SH.Regulation of globin gene expression in erythroid cells.Eur J Biochem,1995,231:271゛281
(1999-01-24 辺後), 為調匳勞
汽了⦿嶄忽匳僥親僥垪 嶄忽亅才匳親寄僥 児粥匳僥冩梢侭匳僥蛍徨伏麗僥忽社嶷泣糞刮片⇧ 臼奨 100005
購囚簡⦿Aγ-帷軌易児咀◉尼強徨◉泣融延◉廬村咀徨GATA-1◉児咀燕器
繁Aγ 姜勣 朕議 冩梢繁Aγ-帷軌易児咀-173 T★C融延斤郡塀恬喘咀徨嚥尼強徨議潤栽式斤尼強徨試來議唹峒。圭隈 寡喘窮咯濡卞個延蛍裂才哽高殆淡烏御児咀鵬扮廬半蛍裂。 潤惚 Aγ-帷軌易児咀-173 T★C融延聞GATA-1嚥融延尼強徨頭粁(-201゛-158)議潤栽週詰阻96%(P<0.01)⇧旺拝聞Oct-1嚥屢揖議DNA頭粁議潤栽週詰55%(P<0.05)。壓MELGM979聾飲嶄⇧融延尼強徨試來頁屎械尼強徨議2蔚(P<0.05)◉壓K562才Hela聾飲嶄⇧融延才屎械尼強徨試來児云屢揖。潤胎 Aγ-尼強徨-173 T★C融延擬崑GATA-1嚥融延尼強徨頭粁潤栽𠮟广受富⇧戻幣壓撹定豚乎廬村咀徨辛嬬頁Aγ-帷軌易児咀議減距陣咀徨◉-173 T★C融延辛嬬叙壓撹定豚碕狼聾飲桟廠嶄嘉嬬奐膿Aγ-帷軌易児咀尼強徨試來。
, http://www.100md.com
嶄夕催 Q512 Q319
Functional Analysis of Human Aγ-globin Gene Promoter
with -173 T★C Mutation
Huang Xiaodong Zhang Junwu# Yang Xuexian Wang Yihong Zhao Hualu
(National Laboratory of Medical Molecular Biology,Institute of Basic of Medical Sciences,CAMS and PUMC,Beijing 100005)
Objective To investigate the effect of the human Aγ-globin gene -173 T★C mutation on the binding of transacting factors to the promoter and the activity of the promoter. Methods Electrophoretic mobility shift assay and transient transfection assay were used in this study. Results The Aγ-globin gene -173 T★C mutation decreased the affinity of GATA-1 to the mutant promoter fragment (-201゛-158)96%(P<0.01) and the binding of Oct-1 to the same DNA fragment 55% (P<0.05).The activity of the mutant promoter was 2-fold (P<0.05) as strong as that of the normal one in MELGM979 cells.The mutant and the normal promoters exhibited basically same activities in K562 and Hela cells. Conclusions The -173 T★C mutation decreased dramatically the binding of GATA-1,implicating that GATA-1 may act as a negative regulator of Aγ-globin gene in adults.The -173 T★C mutation may enhance the activity of Aγ-globin gene promoter in the adult erythroid cell environment.
, 為調匳勞
Key words Aγ-globin gene◉ promoter◉ point mutation◉ transcription factor GATA-1◉ gene expression
繁γ-帷軌易児咀麼勣壓魅故碕聾飲嶄燕器⇧竃伏朔児云購液⇧撹繁碕聾飲嶄魅隅僮碕軌易(HbF,α2γ2)音怎僮碕軌易悳楚議1%⇧遇凖勧來魅隅僮碕軌易隔偬贋壓忝栽屏(HPFH)纂宀議碕聾飲嶄挽醤嗤屢輝互議HbF。HPFH恢伏議圻咀麼勣嗤曾嶽⇧匯頁β-帷軌易児咀関坪議DNA頭粁髪払⇧屈頁γ-帷軌易児咀尼強徨曝議融延。斤HPFH議冩梢嗤廁噐懐苧γ-★β-帷軌易児咀燕器蝕購議字崙。云糞刮冥網-173 T★C融延斤繁Aγ-帷軌易児咀尼強徨孔嬬議唹峒。
1 可創才圭隈
可創 嶷怏嵎腺pNγ才pMγ(蛍艶根屎械式-173 T★C融延議Aγ-帷軌易児咀尼強徨)喇云片更秀。pGL3-Basic墮悶、pSV-β-gal嵎腺、尫崙來坪俳淡、T4 DNA銭俊淡、DNA詔栽淡↔ Klenow頭粁、哽高殆淡(Luc)才β-磯皮滅榠淡(β-Gal)霞協編質歳葎Promega巷望恢瞳。噂*****、Poly[d(I-C)]才屈葬釦滅桓葎Boehringer Mannheim巷望恢瞳。柁互鑓僮碕殆(hemin)才屈遮児冉躾(DMSO)葎Sigma巷望恢瞳。[α-32P]-dGTP葎Amersham巷望恢瞳。DMEM凍劍児、RPMI-1640凍劍児、魅釘僮賠、央路擐扱才N⇧N>-冉遮褒央路擐扱葎Gibco BRL巷望恢瞳。凪万編質葎忽恢蛍裂歓雫。K562、MELGM979才Hela聾飲譲葎云片隠贋。
, 為調匳勞
圭隈
嵎腺議更秀才崙姥⦿蛍艶繍屎械議才-173 T★C融延議Aγ-帷軌易児咀-563゛+14尼強徨頭粁屎﨑嶷怏欺揮嗤Luc烏御児咀議pGL3-Basic謹針臓了泣曝議Kpn↔才Hind‰了泣岻寂⇧資誼燕器嵎腺pNγLuc才pMγLuc(夕1)。參珠叢盾隈戻函嵎腺⇧隼朔喘柁晒錣-粤晒厰凶杢業峠財宣伉隈歓晒[1]。
夕 1 燕器嵎腺pNγLuc才pMγLuc議更秀
Fig 1 Construction of plasmid pNγLuc and pMγLuc
The Nγ and Mγ indicate the normal and mutant Aγ promoter from -563 to +14 respectively
, 為調匳勞
聾飲凍劍⦿侭嗤聾飲譲壓37≧、5%CO2才100%物業議訳周和凍劍。K562才Hela聾飲參根10%魅釘僮賠議DMEM⇧MELGM979聾飲喘根10%魅釘僮賠議RPMI-1640凍劍児凍劍。壓何蛍糞刮嶄⇧K562聾飲壓根嗤25 μmol/L hemin議凍劍児嶄伏海4 d⇧MELGM979聾飲壓根嗤2% DMSO議凍劍児嶄伏海2 d⇧序佩嗾擬蛍晒。
聾飲宰戻函麗議崙姥⦿梓猟孽[2]議圭隈崙姥K562、MELGM979才Hela聾飲宰議戻函麗。梓猟孽[3]議訳周斤宰戻函麗序佩邑裂。霞協宰戻函麗議軌易敵業[4]⇧蛍廾朔贋噐-80≧。
冥寞才蒙呟昇尸DNA議崙姥⦿喘Apa↔才AvaÅ淡盾pNγ才pMγ嵎腺資誼44 bp尼強徨頭粁(-201゛-158)。喘DNA詔栽淡↔ Klenow頭粁才[α-32P]-dGTP挑極温峠炎芝冥寞。掲蒙呟來昇尸DNA葎Poly[d(I-C)]。栽撹議汽全膏宰榠磨将曜諮撹褒全朔恂葎蒙呟來昇尸DNA。冥寞才蒙呟來昇尸DNA議屎吶全宰榠磨乏會需現燕。
, http://www.100md.com
現燕 冥寞式蒙呟來昇尸DNA議屎吶全宰榠磨乏會
Table Sequences of the sense strands of the probes and specific competitors
Name
Sequence (5> to 3>)
Reference
N
CCTTCCCCACACTATCTCAATGCAAATATCTGTCTGAAACGGTC
M
CCTTCCCCACACTATCTCAATGCAAATA
, http://www.100md.com
β-globin GATA-1
CGAGGCCAAGAGATATATCTTAGAGGGAGT
[5]
VH Oct-1
TGCTCATGAATATGCAAATCCTGTGTGTCT
[6]
N.normal probe; M.-173 HPFH mutant probe; asterisk⦿ HPFH mutation site;
underlined nucleotides:Oct-1 binding motif; italic and bold nucleotides:GATA-1 binding motif
, http://www.100md.com
窮咯濡卞個延蛍裂⦿20 μl議DNA-軌易嵎悶翌潤栽郡哘悶狼嶄茅根嗤10 μg聾飲宰戻函麗才2 μg Poly[d(I-C)]翌凪万撹蛍嚥猟孽[3]烏御議屢揖。郡哘詞栽麗噐30≧剣圄5 min⇧壅紗秘冥寞(埃1 ng)賜紗秘冥寞式蒙呟昇尸DNA議詞栽麗⇧隼朔噐30≧写偬剣圄15 min。郡哘恢麗梓猟孽[3]議訳周序佩窮咯⇧噐-70≧序佩慧符徭𠮟唹。喘廾嗤Gelscan XL 2.1罷周議Pharmacia LKB UltroScan XL爾高畜業膝宙卩霞協嵳藻揮議高畜業。乎糞刮嶷鹸4肝。
鵬扮廬半蛍裂⦿梓猟孽[7]議圭隈廬半K562才MELGM979聾飲。繍10 μg pNγLuc賜pMγLuc、2 μg pSV-β-gal才30 μg沜娼DNA擬秘2〜107倖聾飲嶄。喘宋磨呼-DNA慌柿牛隈[1]繍8 μg pNγLuc賜pMγLuc才1.6 μg pSV-β-gal廬半欺5〜105倖Hela聾飲嶄。pSV-β-gal恬葎坪斤孚⇧喘參丕屎廬半丼楕。廬半議聾飲凍劍48 h朔⇧梓孚編質歳荷恬傍苧序佩β-gal式Luc議試來霞協。糞刮嶷鹸3肝⇧耽肝恂3倖峠佩廬半。參pNγLuc議Luc試來峙恂葎児彈1.0⇧繍pMγLuc議Luc試來峙嚥岻序佩曳熟⇧箔竃屢斤曳峙⇧喘參燕幣融延嚥屎械尼強徨議試來曳。
, http://www.100md.com
由柴僥侃尖⦿糞刮方象參譲峙\炎彈餓燕幣⇧怏寂𠮟广來餓呟曳熟寡喘t殊刮。
2 潤惚
融延斤郡塀恬喘咀徨嚥尼強徨潤栽議唹峒 窮咯濡卞個延蛍裂(夕2)燕苧⦿K562、MELGM979才Hela聾飲議宰戻函麗嚥冥寞潤栽朔譲辛侘撹嵳藻揮A⇧融延冥寞議A揮曳屎械冥寞A揮議高畜業週詰(55\4)%(n=4,P<0.05)◉K562才MELGM979聾飲議宰戻函麗珊辛參嚥冥寞潤栽侘撹嵳藻揮B⇧融延冥寞議B揮曳屎械冥寞B揮議高畜業週詰(96\3.5)%(n=4,P<0.01)。蒙呟來昇尸潤栽糞刮屬苧⇧嵳藻揮A頁Oct-1嚥冥寞潤栽朔侘撹議⇧遇嵳藻揮B頁GATA-1嚥冥寞潤栽朔侘撹議。
夕 2 屎械式融延冥寞嚥K562、MELGM979式Hela聾飲宰戻函麗潤栽議窮咯濡卞個延蛍裂
, http://www.100md.com
Fig 2 Electrophoretic mobility shift assay of the normal and mutant probes using nuclear extracts from K562,MELGM979 and Hela cells
N.normal AvaÅ-Apa↔ fragment of Aγ-promoter(-201゛-158);
M.-173 HPFH mutant counterpart of N; MEL.MELGM979; (+).nuclear extracts from the hemin/DMSO-induced cells; A. band retarded by binding of Oct-1; B.band retarded by binding of GATA-1
融延斤尼強徨試來議唹峒 Luc試來霞協𠮟幣⇧壓DMSO嗾擬議才隆嗾擬議MELGM979聾飲嶄⇧融延Aγ-児咀尼強徨議試來蛍艶屢輝噐屎械尼強徨試來議(2.01\0.25)蔚(n=3,P<0.05)才(1.96\0.04)蔚(n=3,P<0.05)◉壓Hela聾飲才hemin嗾擬才隆嗾擬議K562聾飲嶄⇧融延Aγ-児咀尼強徨議試來蛍艶屢輝噐屎械尼強徨試來議(0.97\0.03)、(0.98\0.03)才(0.99\0.02)蔚⇧嚥屎械尼強徨試來議餓呟譲涙𠮟广來吭吶(n=3,P>0.05)。
, http://www.100md.com
3 網胎
児咀議廬村距陣麼勣頁宥狛DNA嚥軌易嵎參式軌易嵎嚥軌易嵎岻寂議屢札恬喘糞孖議。Aγ-帷軌易児咀尼強徨議-190゛-170曝囃淫凄曾倖GATA-1潤栽了泣才匯倖了噐屈宀岻寂議Oct-1潤栽了泣⇧凪嶄Oct-1潤栽了泣嚥和嗄議GATA-1潤栽了泣何蛍嶷京。云冩梢潤惚燕苧嚥Aγ-児咀壓撹繁豚試埔燕器屢購議-173 T★C融延苧𠮟週詰阻GATA-1嚥緩融延侭壓議尼強徨曝囃議潤栽。Berry吉[8]奚烏御嚥HPFH燕侏屢購議Aγ-尼強徨-117 G★A融延匆週詰阻GATA-1嚥尼強徨議潤栽⇧旺喇緩邪譜潤栽欺-117曝囃議GATA-1辛嬬斤撹繁豚議γ-帷軌易児咀軟減距陣恬喘。永宀容霞潤栽欺-173曝囃議GATA-1斤撹繁豚議γ-帷軌易児咀辛嬬匆揖劔軟減距陣恬喘。參吏議烏御屬苧壓俯謹秤趨和GATA-1頁廬村議試晒咀徨[9]⇧徽頁緩咀徨議恬喘辛嬬頁鹸墫議⇧賜俯嚥醤悶議DNA潤栽了泣式窟圄竣粁嗤購⇧遇拝辛嬬卆正噐嚥凪万廬村咀徨議屢札恬喘。
, 為調匳勞
Luc試來霞協潤惚𠮟幣⇧壓MELGM979嶄-173 T★C融延尼強徨試來屢輝噐屎械尼強徨議2蔚⇧遇壓K562才掲碕狼Hela聾飲嶄融延才屎械尼強徨試來涙餓艶。K562才MELGM979譲葎碕狼易僮押聾飲⇧念宀嬬戻工冬魅豚才魅隅豚碕狼聾飲桟廠⇧遇朔宀戻工冬魅豚才撹定豚碕狼聾飲桟廠。喇緩燕苧⇧-173 T★C融延辛嬬叙壓撹定豚碕狼聾飲桟廠嶄嘉嬬奐膿Aγ-帷軌易児咀尼強徨試來。徽峙誼廣吭議頁⇧軸聞壓MELGM979聾飲嶄⇧-173 T★C融延擬崑Aγ-尼強徨試來議奐紗(1蔚)匆垓音式悶坪(>20蔚)議互⇧咀緩叙喘匯倖燕器嵎腺序佩議鵬扮燕器珊音嬬郡啌悶坪議寔糞彜趨。云冩梢潤惚葎-173 T★C融延唹峒Aγ-帷軌易児咀窟圄蝕購序殻議字崙戻工阻匯協㞍沫⇧隼遇凪鳩俳字崙賓棋序匯化冩梢。
歌深猟孽
1 Sambrook J,Fritsh EF,Maniatis T.Molecular cloning:A laboratory manual.2nd ed.New York:Cold Spring Harbor Laboratory Press,1989
, 為調匳勞
2 Dignam JD,Lebovitz RM,Roeder RG.Accurate transcription initiation by RNA polymerase Å in a soluble extract from isolated mammalian nuclei.Nucleic Acid Res,1983,11:1475゛1489
3 Gumucio DL,Rood KL,Gray TA,et al.Nuclear proteins that bind the human γ-globin gene promoter:alterations in binding produced by point mutations associated with hereditary persistence of fetal hemoglobin.Mol Cell Biol,1988,8:5310゛5322
4 Braford MM.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding.Anal Biochem,1976,72:248゛254
, 為調匳勞
5 doBore E,Antonion M,Mignotte V,et al.The human β-globin promoter,nuclear protein factors and erythroid specific induction of transcription.EMBO J,1988,7:4203゛4212
6 Ballard DW,Bothwell A.Mutational analysis of the immunoglobulin heavy chain promoter region.Proc Natl Acad Sci USA,1986,83:9626゛9630
7 嫖拭冷⇧孕凩埴⇧伶酵椰⇧吉.勸伏侏式尼強徨曝融延議HPFHAγ児咀壓MEL聾飲嶄議燕器.伏麗晒僥墫崗⇧1995⇧11⦿343゛347
8 Berry M,Grosveld F,Dillon N.A single point mutation is the cause of the Greek form of hereditary persistence of fetal haemoglobin.Nature,1992,358:499゛502
9 Orkin SH.Regulation of globin gene expression in erythroid cells.Eur J Biochem,1995,231:271゛281
(1999-01-24 辺後), 為調匳勞