小麦黄花叶病毒和小麦梭条斑花叶病毒的生物学和分子生物学研究
作者:陈炯 程晔 陈剑平
单位:陈炯(浙江大学生物科学与技术系,杭州 310027);程晔 陈剑平(浙江省农业科学院真菌传植物病毒实验室,杭州 310021)
关键词:小麦黄花叶病毒;小麦梭条斑花叶病毒;流行病学;基因组结构
中国病毒学000201 中图分类号:S432.41 文献标识码:A
文章编号:1003-5125(2000)02-0097-09
Biological and Molecular Biological Characterization of Wheat Yellow Mosaic and Wheat Spindle Streak Mosaic Bymoviruses
, 百拇医药
CHEN Jiong
(Department of Biological Science and Technology,Zhejiang University,Hangzhou 310027,China)
CHENG Ye,CHEN Jian-ping
(Virology Laboratory,Zhejiang Academy of Agricultural Sciences,Hangzhou 310021,China)
Key words:Wheat yellow mosaic bymovirus; Wheat spindle streak mosaic bymovirus; Epidemiology; Genome organization
, http://www.100md.com 线状土传小麦花叶病毒病在欧洲、亚洲和北美洲等地均有发生。这类病毒由根肿菌纲的禾谷多粘菌(Polymyxa graminis)传播,并归类于马铃薯Y病毒科(Potyviridae)的大麦黄花叶病毒属(Bymovirus)。
日本的Sawada(1927)[1]首次发现并描述小麦花叶病害,随后的研究表明病原是土传的。Inouye(1969)[2]证明该病害的病原是小麦黄花叶病毒(wheat yellow mosaic virus,WYMV)。Slykhuis(1960)[3]首次在加拿大安大略省的冬小麦上发现并描述小麦梭条斑花叶病害。Slykhuis和Polak(1969)[4]证明该病害是由小麦梭条斑花叶病毒(wheat spindle streak mosaic virus,WSSMV)引起的。这两个名字都曾被用于我国和欧洲分离物,是我国冬麦区包括陕西、四川、湖北、山东、河南、安徽、江苏和浙江等省的重要病害。在德国,曾报道在黑麦上分离到WYMV。由于WSSMV和WYMV症状和细胞病理学、血清学、病毒粒子形态学、基因组和外壳蛋白大小以及小麦品种反应的相似性,Usugi和Saito(1979)[5]建议WSSMV为WYMV的一个株系。此后,Sohn等(1994)[6]报道了WSSMV法国分离物RNA1 3′-端4kb序列,于嘉林等(1995)[7]报道了一个中国河南分离物RNA1 3′-末端的891个核苷酸序列,发现与法国分离物仅有69.9%的序列同源性。最近,Namba等(1998)[8]报道了日本WYMV基因组全长序列,与法国分离物相比,也仅有69.8%的同源性,从而表明它们是不同的病毒。以下阐述这两种病毒的生物学和分子生物学特性。
, http://www.100md.com
1 生物学和生态学
1.1 宿主范围和症状
两种病毒在自然界都仅侵染普通小麦(Triticum aestivum)和硬粒小麦(T.durum)。曾有报道说WYMV在德国的黑麦(Secale cereale)上也有发现[9],然而,最近的研究表明该分离物是WSSMV[10]。在实验室条件下,两种病毒均能摩擦接种小麦和黑麦。
WYMV侵染的小麦在2月中旬显症,嫩叶上呈现褪绿条纹或黄花叶症状,在老叶上也常出现坏死斑。3~5月间气温升高后,花叶症状逐渐消失,新叶无症,但是植株分蘖减少。通常感病植株麦穗短小,发育不全。WSSMV症状通常为短褪绿或坏死斑点及梭条状条纹,引起小麦分蘖减少及矮化。温度是该类病害发展及症状表现的主要决定因素。
症状表现由于不同小麦品种而有所差别。通常感病品种症状严重,而抗病品种则无症或症状轻微。两种病害的田间症状都是田块呈现浅绿色到黄色,很容易和肥力缺乏引起的作物田间症状相混淆。
, 百拇医药
1.2 地理分布
土传线状小麦花叶病害在亚洲、欧洲和北美洲普遍发生,主要分布于北半球的冬小麦栽种区。已证实其存在的地区为:中国的安徽、河南、江苏、湖北、陕西、四川、山东和浙江等省,印度,日本的北海道、本州、九州、琉球和四国,加拿大的安大略省,美国的阿拉巴马、阿肯萨斯、乔治亚、伊利诺斯、印地安那、肯萨斯、肯特基、马里兰、密歇根、内布拉斯加、纽约、俄克拉哈马、宾西法尼亚和弗吉尼亚等州,意大利,法国和德国等。
通过不同地区分离物的基因组研究证实,欧洲和北美洲分离物是一个基因组同源的种,彼此间非常相似(93.5%~99.5%);中国和日本分离物是另一个基因组同源的种群(96.9%~99.0%);与欧美分离物具有很大的差异(核苷酸序列同源性67.7%~77.7%)。因此北美和欧洲分离物应当统一为WSSMV,而亚洲分离物应当为WYMV[10]。
1.3 病害传播
, http://www.100md.com
1.3.1 介体
禾谷多粘菌为根肿菌纲成员。Ledingham(1939)在加拿大安略省的小麦上首次发现该菌,通过9年观察,他对禾谷多粘菌的形态学和生活史作了详细的描述[11]。陈剑平等(1992,1993,1998)[12~14]和Littlefield等(1997)[15]对禾谷多粘菌超微结构作了详尽的研究。Castlebury和Domier(1998)[16]研究了根肿菌纲代表成员甘蓝根肿菌(Plasmodiophora brassicae)的进化史,他们在分析了核糖体小亚单位RNA基因(SSU rDNA)的序列数据后发现,它和对照的生物亲缘性都不高,但和原生动物的亲缘性要比它和真菌或是粘菌的亲缘性更高。禾谷多粘菌的宿主范围很广,包括禾谷类的大麦、小麦和燕麦,一些分离物还能侵染双子叶植物。
1.3.2 病毒-介体关系
, 百拇医药
Slykhuis和Barr(1978)[17]提出WSSMV是由禾谷多粘菌传播。他们用禾谷多粘菌接种小麦植株的根系,和(或)在植物叶片上摩擦接种病毒,然后将一些无病幼苗和它们栽种在一起,结果表明只有在WSSMV和禾谷多粘菌都存在的情况下,幼苗上才会发生病害,且该病害不能由其他测试的土栖真菌孢子传播。土壤中WSSMV的侵染性经52.5℃加热30min丧失,湿土或干土的侵染性在温室内或户外都可保持5年。
对于WYMV病害传播的研究不多。Ikata和Kawai(1940)[18]调查了WYMV病害的生态学,揭示该病害的病原是通过土壤传播的。带毒的土壤在户外放置5年或室内4年后丧失侵染性。因为WYMV和大麦黄花叶病毒(barley yellow mosaic virus,BaYMV)及WSSMV非常相似,故一般认为病毒存在于禾谷多粘菌体内以持续性方式传播。
1.3.3 病害流行学
, 百拇医药
秋季适当降雨和较低温度(4~13℃,最适为10℃)、冬季寒冷以及春季延续的低温有利于病害的发生和症状表现。连作感病小麦品种会导致该类病害的流行,并且病症加重,休耕能消除或降低土壤的侵染性。温度是病害发展及症状表现的主要决定因素[18,19]。
WSSMV及WYMV侵染冬小麦的最适温度为15℃(变动范围可在5~17℃,但不能超过20℃)。在适宜的温度及充足的土壤湿度条件下,小麦播种后4~10d病毒侵染就会发生,10~20d侵染达到高峰[18~20]。在北温带小麦栽种区,大约是在播种后10~40d,田间的病毒就随着禾谷多粘菌的游动孢子开始侵染小麦。播种30d后,用ELISA方法可以在冬小麦根部检测到病毒,60d后,就能在嫩叶上检测到病毒。
病毒侵染的植株数以及根组织中的病毒含量在秋冬季增加,嫩叶中的病毒含量在秋末、冬季和早春增加。通常这段时间的气温都低于5℃,表明病毒的二次侵染、复制、细胞-细胞间的运动以及系统性的运动都只在很低的温度条件下发生[20~22]。
, 百拇医药
这类病毒从根到叶的运动相当缓慢,约需30~40d,而从叶到根的运动仅需要5d。病毒从根到叶的运动之所以缓慢,可能取决于韧皮部对病毒的吸收情况[23]。
虽然秋冬季时ELISA法已能检测到植株中病毒的存在,但此时小麦植株是无症的[20~22]。冬末早春时分,温度常变动在5~13℃,植株经春化后重新生长,新叶才逐渐表现出症状。病土接种的植株,在合适温度下,通常需60d或更长的潜伏期才能表现出症状;即便是用病毒摩擦接种幼苗,症状表现也需要经过30~40d时间[24,25]。
用以下一些处理方法可以使潜伏期缩短几周,并使表现症状植株的百分数增加:1)交替的冰冻和融冻;2)培养温度在5~17℃范围内变动;3)严重冰冻(-17℃)损害后再生长;4)剪除植株嫩叶后再生长。除方法2外,其余方法都是对根(直接)和(或)叶片(直接或间接)的损害,推测可能是由于暂时性地改变了植株韧皮部结构,病毒更容易进入,从而导致潜伏期的缩短[23]。
, http://www.100md.com
1.3.4 经济影响
WSSMV和WYMV的危害主要表现为小麦分蘖减少、生长受阻、产量减少和质量下降。
在加拿大安大略省,WSSMV侵染引起的冬小麦产量损失一般在3%~59%之间。在美国北部、中部和东北部地区,WSSMV的侵染常导致24%~64%的减产。欧洲WSSMV通常是和土传小麦花叶病毒(soil-borne wheat mosaic virus,SBWMV)混合侵染小麦的,引起的作物产量损失也很严重。中国WYMV病害分布区域包括北部冬小麦区的胶东沿海区,黄淮冬小麦区的大部,长江中、下游冬小麦区的部分和西南冬小麦区的四川盆地区等小麦生态区。据我们1998年春天沿312国道从南京到河南潢川的调查,大约有20%~30%的小麦发病。如河南潢川县,WYMV常年发病面积在10000hm2;1991年大发生,发病面积23333hm2,3333hm2绝收;1970~1998年累计发病面积达206666hm2。一般病田小麦减产10%~30%,重病田减产可高达50%~70%。山东胶东半岛的小麦花叶病害是WYMV和中国小麦花叶病毒(Chinese wheat mosaic virus,CWMV)混合侵染引起的,危害更为严重,感病品种常常整片坏死[26]。在日本,WYMV可引起小麦产量的严重损失,高感病和轻微感病品种间存在35%的产量差异。
, http://www.100md.com
1.3.5 病害防治
由于这类病毒传播介体的休眠孢子抗逆性很强,化学试剂和杀菌剂的使用在经济和生态学上都是不可行的,休耕或轮作可以起到一定的防治作用,但是这类病害的控制主要靠栽种抗病品种。由于目前无法筛选到抗禾谷多粘菌的小麦品种,所以今后的育种仍有赖于病毒基因介导的抗性。浙江省农科院病毒室和德国马普所、中国农科院植保所及英国洛桑试验站合作,应用基因工程改良小麦,使小麦对这类病害产生抗性。目前全长病毒外壳蛋白和截短的NIb基因小麦表达载体已经构建完成,小麦转基因工作正在进行之中。
2 分子生物学
2.1 病毒粒子性质
WYMV和WSSMV均为弯曲的线状病毒粒子。WSSMV粒子典型长度为275~300nm和600~625nm,直径为12.8nm。提纯的线状病毒长度常常超过2000nm,可能是由于病毒粒子两端线性化聚集引起[4]。WYMV粒子长度典型分布峰为275~300nm和575~600nm,直径13~14nm[2]。
, 百拇医药
两种病毒都能在宿主植物中产生细胞质内含体。在WYMV侵染的小麦植株叶片细胞中已发现有复合膜状体、风轮体和圆柱状体的存在。这些内含体通常成簇出现并在原生质体中占据了很大的空间。病毒状粒子散布在原生质体中,或与小的结晶体及风轮状内含体结合[2,27]。WSSMV侵染的小麦细胞病理学和WYMV相似,膜状体通常是有序排列的。细胞质的某些部位存在很多风轮状内含体,与WYMV侵染细胞中的情况很相似。病毒状粒子散布或聚集在细胞质中,或结合有一些小的结晶体。粒子也沿着风轮状内含体的长轴方向并行排列[27,28]。
WYMV、WSSMV和BaYMV血清学相关。免疫电镜研究表明,BaYMV和WYMV抗血清反应较强,与美国WSSMV抗血清反应较弱,而与大麦和性花叶病毒(barley mild mosaic virus,BaMMV)或SBWMV抗血清不反应[10]。
2.2 基因组结构及编码的蛋白
, 百拇医药
两种病毒均具有两条单股、正链RNA基因组。WSSMV RNAs的分子量分别为2.6×106Da和1.4×106Da;WYMV RNAs的分子量分别为2.6×106Da和1.5×106Da。由WSSMV和WYMV外壳蛋白基因的核苷酸序列推测的外壳蛋白大小(CP)分别为31.9kDa和32kDa,和SDS-PAGE分析结果几乎相同[6,8,29]。WSSMV和WYMV的基因组结构相似,但同源性较低(图1和表1)。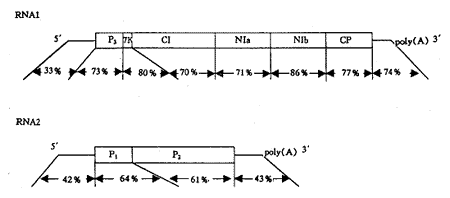
图1 WSSMV-Fr和WYMV-T基因组结构图示及不同区域的同源性
Fig.1 Genomic organization and homology of WSSMV-Fr and WYMV-T
, 百拇医药
表1 WYMV-T和WSSMV-Fr的基因组结构及其编码蛋白
Table 1 Genomic organization and induced proteins of WYMV-T and WSSMV-Fr
WYMV-T
WSSMV-Fr
核苷酸
蛋白质
核苷酸
蛋白质
位置
大小
位置
, 百拇医药
大小
Mr(kD)
位置
大小
位置
大小
Mr(kD)
RNA1
5′-UTR
1~162
162
1~169
169
, http://www.100md.com
启始密
163~165
170~172
码子
P3
163~1143
981
1~327
327
36
170~1099
930
1~310
, http://www.100md.com
310
35
7kDa
1144~1341
198
328~393
66
7
1100~1297
198
311~376
66
7
, http://www.100md.com
CI
1342~3318
1977
394~1052
659
73
1298~3274
1977
377~1035
659
73
NIa
3319~4911
, 百拇医药
1593
1053~1583
531
60
3275~4970
1596
1036~1567
532
60
NIb
4912~6495
1584
1084~2111
, http://www.100md.com
528
59
4971~6454
1584
1568~2095
528
59
CP
6496~7374
879
2112~2404
293
32
, 百拇医药
6455~7336
882
2096~2389
294
32
ORF
163~7374
7212
1~2404
2404
269
170~7336
7167
, http://www.100md.com
1~2389
2389
267
终止子
7375~7377
7337~7339
3′-UTR
7378~7636
259
7340~7570
231
RNA2
, 百拇医药 168
5′-UTR
1~168
1~170
170
启始密
169~171
171~173
码子
P1
169~933
765
1~55
, 百拇医药
255
28
171~935
765
1~255
255
28
P2
934~2880
1947
256~904
649
73
, http://www.100md.com
936~2870
1935
256~900
635
71
ORF
169~2880
2712
1~904
904
101
171~2870
2700
, 百拇医药
1~900
900
100
终止子
2881~2883
2871~2873
3′-UTR
2884~3659
776
2874~4073
1200
WYMV-T(WYMV日本典型分离物)RNA1和RNA2分别由7636和3659nt组成[不包括3′-poly(A)尾]。计算机分析揭示,RNA1和RNA2分别具有一个长的开放阅读框架(ORF1和ORF2)。ORF1长7212nt,起始于nt 163~165处的第一个AUG,终止于nt 7375~7377处的UAA密码子。其后是长为259nt的3′-UTR。ORF1编码一个2404个氨基酸(aa)的蛋白质,分子量269kDa。推测269kDa蛋白经切割后产生6个蛋白,包括C端的CP和5个非结构蛋白。CP由293aa组成,分子量32kDa。NIb蛋白具有RNA依赖性的RNA聚合酶活性位点的保守模式SGX3TX3NT X18~37GDD(1915~1965aa)。NIa-Pro负责269kDa蛋白的切割。NIa-VPg含有一个Tyr,可能是起结合病毒RNA5′-末端的作用。CI蛋白具有NTP结合模式GXXGXGKS(486~583aa),不同处是第四个氨基酸G被V替换,被认为具有解螺旋酶的活性。P3蛋白具有锌指结构。7kDa蛋白功能仍不清楚。ORF2长为2712nt,起始于nt 169~171处的第一个AUG密码子,终止于nt 2881~2883处的UAG密码子,推测编码一个904aa、分子量101kDa的蛋白。101kDa蛋白在254~256aa处含有一个VGS三氨基酸切割位点,通过切割可产生两个大小分别为28kDa(P1)和73kDa(P2)的蛋白。P1蛋白含有蛋白酶结构域GYCY(141~144aa),并在215aa处有一个切割活性所必需的组氨酸残基,该蛋白和马铃薯Y病毒科其他成员的HC-Pro蛋白结构类似。P2蛋白含有一个假的CP结构域(319~398aa)和一个富含亮氨酸的结构域(595~777aa)。与马铃薯Y病毒科其他病毒的P2蛋白以及Furovirus的CP-readthrough(RT)蛋白具有一定的同源性。WYMV RNA1和RNA2的5′-UTR长度相似(分别为162和168nt),且具有相当的序列同源性(65%)。5′-UTR可能在病毒包装、复制及翻译中起着重要的作用。WYMV RNA1和2的3′-UTR在长度(分别为259和767nt)和序列上都具有很大差异[8,10]。
, http://www.100md.com
WSSMV-Fr(WSSMV法国分离物)RNA1和RNA2分别长为7570和4073核苷酸[不包括3′-poly(A)尾]。基因组结构和WYMV-T相似。RNA1 ORF1长7167核苷酸,起始于nt 170~172处的第一个AUG,终止于nt 7337~7339处的UAA密码子,编码一个2389aa的蛋白质,分子量266.5kDa;3′-UTR长231nt。ORF2长2700nt,起始于nt 171~173处的第一个AUG密码子,终止于nt 2871~2873处的UAG密码子,推测编码一个900aa、分子量99.8kDa的蛋白。两个聚合蛋白的结构和WYMV类似。RNA1和RNA2的5′-UTR长度相似(分别为169和170nt),3′-UTR在长度(分别为231和1200nt)和序列上都具有很大差异(表1)[6,30]。
2.3 分子进化和株系分化
WSSMV和WYMV外壳蛋白区域的序列同源性与确立的马铃薯Y病毒科不同种的划分标准是相符的,但较高的同源性也揭示WSSMV、WYMV和BaYMV是很相似的病毒。对于外壳蛋白区域核苷酸序列的相关性研究表明了这些病毒分离物间的近缘关系(图2)[10]。
, http://www.100md.com
1991年,在日本北海道地区Eniwa城发现一个有别于WYMV-T的株系,被命名为WYMV-H。在1991~1994年,Kusume等研究表明WYMV-T、WYMV-H和WSSMV在不同小麦品种上的致病性存在明显差异。WYMV-T能侵染关东地区的主要秋播小麦品种Norin 61,而WYMV-H不能;北海道地区的两种主要秋播小麦品种Chihokukomugi和Horoshirilomugi仅被WYMV-H侵染;且WYMV-H不侵染对WYMV-T高感病的品种Hatakedakomugi。ELISA结果也进一步表明了WYMV-T、WYMV-H和WSSMV之间的血清学差异,WYMV-H株系与WYMV-T在血清学上关系更近[3]。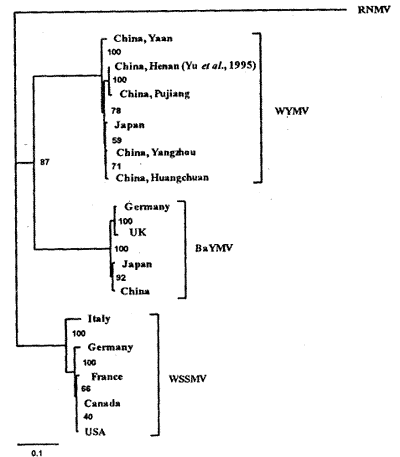
图2 使用GCG程序PILEUP、DISTANCES和GROWTREE对WYMV、WSSMV和BaYMV各分离物的外壳蛋白核苷酸序列的分析,水稻坏死花叶病毒(rice necrosis mosaic virus,RNMV)作为一个根植的外围集团用TREEVIEW程序表示各分离物间的相关性。比例尺代表每100个碱基中不同碱基的数目[10]。
, http://www.100md.com
Fig.2 Phylogram of the coat protein nucleic acid sequences of isolates of WYMV,WSSMV and BaYMV prepared from the GCG programs PILEUP,DISTANCES and GROWTREE and displayed in TREEVIEW with RNMV as the rooted outgroup.The scale bar shows the number of substitutions per 100 bases.
我国WYMV分离物的部分序列测定表明它们的同源性较高,但初步田间试验表明不同试点的品种反应存在差异,因而可能存在不同于日本分离物的株系,有待进一步证实。
对于WSSMV的株系分化情况,至今尚无报道。
基金项目:863资助项目(101-04-01-05);浙江省重大科技项目(961106129);浙江省青年科技人才专项资金(RC9604)
, 百拇医药
作者简介:陈炯(1975年-),男,汉,浙江绍兴人,硕士,1999年入学读博士学位。主要从事植物病毒学研究。
参考文献
[1]Sawada E.Wheat yellow mosaic prevention[J].Journal of Plant Protection (Byochugai-Zasshi),1927,14:444~449
[2]Inouye T.Viral pathogen of the wheat yellow mosaic disease[J].Nogaku Kenkyu,1969,53:61~68
[3]Slykhuis JT.Evidence of soil-borne mosaic of wheat in Ontario[J].Canadian Plant Disease Survey,1960,40:43
, 百拇医药
[4]Slykhuis JT.Verification of wheat spindle streak mosaic virus in Ontario[J].Canadian Plant Disease Survey,1969,49:108~111
[5]Usugi T,Saito Y.Relationship between wheat yellow mosaic virus and wheat spindle streak mosaic virus[J].Annals of the Phytopathological Society of Japan,1979,45:397~400
[6]Sohn A,Schenk P,Signoret PA,et al.Sequence analysis of the 3′ terminal half of RNA1 of wheat spindle streak mosaic virus[J].Archives of Virology,1994,135:279~292
, http://www.100md.com
[7]于嘉林,晏立英,冯继东,等.一种中国发生的真菌传小麦花叶病毒RNA-1 3′末端核苷酸序列分析[J].病毒学报.1995,11(5):248~254
[8]Namba S,Kashiwazaki S,Lu X,et al.Complete nucleotide sequence of wheat yellow mosaic bymovirus genomic RNAs[J].Archives of Virology,1998,143,631~643
[9]Huth W,Lesemann D-E.Fungus-transmitted soil-borne viruses on rye in Germany[J].Journal of Plant Diseases and Protection,1993,103:120~124
[10]Jiong Chen,Anke Sohn,Jianping Chen,et al.Molecular comparisons amongst wheat bymovirus isolates from Asia,North America and Europe[J].(in submission)
, 百拇医药
[11]Leddingham GA.Studies on Polymyxa graminis,n.gen.n.sp.,a plasmodiophoraceous root parasite of wheat[J].Canadian Journal of Research C,1939,17:38~51
[12]陈剑平,Adams M J.禾谷多粘菌原质因和游动孢子囊的超微结构[J].浙江农业学报,1993,5:71~74
[13]陈剑平,Adams M J.禾谷多粘菌游动孢子超微结构[J].真菌学报,1992,11(3):216~220
[14]Chen JP,Wang Z,Hong J,et al.Ultrastructural studies of resting spore development in Polymyxa graminis[J].Mycol Res,1998,102(6):687-691
, 百拇医药
[15]Littlefield LJ,Delfosse P,Whallon JH,et al.Anatomy of sporosori of Polymyxa graminis,the vector of Indian peanut clump virus,in roots of Sorghum bicolor[J].Canadian Journal of Plant Pathology,1997,19:281~288
[16]Castlebury LA,Domier LL.Small subunit ribosomal RNA gene phylogeny of Plasmodiophora brassicae[J].Mycologia,1998,90:102~107
[17]Slykhuis JT,Barr DJS.Confirmation of Polymyxa graminis as a vector of wheat spindle streak mosaic virus[J].Phytopathology,1978,68:639~643
, http://www.100md.com
[18]Ikata S,Kawai I.Studies on wheat yellow mosaic disease[J].Noji-kairyo-shiryo,Ministry of Agriculture and Forestry,1940,154:1~123
[19]Slykhuis JT.Factors determining the development of wheat spindle streak mosaic caused by a soil-borne virus in Ontario[J].Phytopathology,1970,60:319~331
[20]Ohto Y,Naito S.Propagation of wheat yellow mosaic virus in winter wheat under low temperature conditions[J].Annals of the Phytopathological Society of Japan,1997,63:361~365
, http://www.100md.com
[21]Cunfer BM,Demski JW,Bays DC.Reduction in plant development,yield,and grainquality associated with wheat spindle streak mosaic virus[J].Phytopathology,1988,78:19~204
[22]Carroll JE[J],Bergstrom GC,Gray SM.Dynamics of wheat spindle streak mosaic bymovirus in winter wheat[J].European Journal of Plant Pathology,1997,103:313~321
[23]Schenck PM,Antoniw JF,Batista MdeF,et al.Movement of barley mild mosaic and baley yellow mosaic virus in leaves and roots of barley[J].Annals of Applied Biology,1995a,126:291~305
, http://www.100md.com
[24]Slykhuis JT.Factors critical to mechanical transmissibility of wheat spindle streak mosaic virus[J].Phytopathology,1975a,65:582~584
[25]Usugi T.Epidemiology and management in Japan of soil-borne cereal mosaic viruses with filamentous particles.In:Cooper JI,Asher MJC eds.Developments in Applied Biology 2,1988,Viruses with Fungal Vectors[M].Wellesbourne,UK:Association of Applied Biologists,1988.213~225
[26]Rong Ye,Tao Zheng,Jianping Chen,et al.Characterization and partial sequence of a new Furovirus of wheat in China[J].Plant Pathology,(Accepted)
, 百拇医药
[27]Hibino H,Usugi T,Saito Y.Comparative electron microscopy of inclusions associated with five soil-borne filamentous viruses of cereals[J].Ann Phytopath Soc Japan,1981,47:510~519
[28]Hooper GR,Wiese MV.Cytoplasmic inclusions in wheat affected by wheat spindle streak mosaic[J].Virology,1972,47:664~672
[29]Usugi T,Kashiwazaki S,Omura T,et al.Some properties of nucleic acids and coat proteins of soil-borne filamentous viruses[J].Ann Phytopath Soc Japan,1989,55:26~31
, 百拇医药
[30]Sohn A,Leclair S,Steinbiss H-H.Analysis of RNA1 of wheat spindle streak mosaic bymovirus (WSSMV)[C].In:Sherwood JL,Rush CM eds.Proceedings of the Third Symposium of the Internationsal Working Group on Plant Viruses with Fungal Vectors,Dundee.American Society of Sugaar Beet Technologists.USA:Denver,Co.1996.45~48
[31]Kusume T,Tamada T,Hattor H,et al.Identification of a new wheat yellow mosaic virus strain with specific pathogenicity towards major wheat cultivars grown in Hokkaido[J].Ann Phytopath Soc Japan,1997,63:107~109
收稿日期:1999-01-25
修回日期:1999-05-05, http://www.100md.com
单位:陈炯(浙江大学生物科学与技术系,杭州 310027);程晔 陈剑平(浙江省农业科学院真菌传植物病毒实验室,杭州 310021)
关键词:小麦黄花叶病毒;小麦梭条斑花叶病毒;流行病学;基因组结构
中国病毒学000201 中图分类号:S432.41 文献标识码:A
文章编号:1003-5125(2000)02-0097-09
Biological and Molecular Biological Characterization of Wheat Yellow Mosaic and Wheat Spindle Streak Mosaic Bymoviruses
, 百拇医药
CHEN Jiong
(Department of Biological Science and Technology,Zhejiang University,Hangzhou 310027,China)
CHENG Ye,CHEN Jian-ping
(Virology Laboratory,Zhejiang Academy of Agricultural Sciences,Hangzhou 310021,China)
Key words:Wheat yellow mosaic bymovirus; Wheat spindle streak mosaic bymovirus; Epidemiology; Genome organization
, http://www.100md.com 线状土传小麦花叶病毒病在欧洲、亚洲和北美洲等地均有发生。这类病毒由根肿菌纲的禾谷多粘菌(Polymyxa graminis)传播,并归类于马铃薯Y病毒科(Potyviridae)的大麦黄花叶病毒属(Bymovirus)。
日本的Sawada(1927)[1]首次发现并描述小麦花叶病害,随后的研究表明病原是土传的。Inouye(1969)[2]证明该病害的病原是小麦黄花叶病毒(wheat yellow mosaic virus,WYMV)。Slykhuis(1960)[3]首次在加拿大安大略省的冬小麦上发现并描述小麦梭条斑花叶病害。Slykhuis和Polak(1969)[4]证明该病害是由小麦梭条斑花叶病毒(wheat spindle streak mosaic virus,WSSMV)引起的。这两个名字都曾被用于我国和欧洲分离物,是我国冬麦区包括陕西、四川、湖北、山东、河南、安徽、江苏和浙江等省的重要病害。在德国,曾报道在黑麦上分离到WYMV。由于WSSMV和WYMV症状和细胞病理学、血清学、病毒粒子形态学、基因组和外壳蛋白大小以及小麦品种反应的相似性,Usugi和Saito(1979)[5]建议WSSMV为WYMV的一个株系。此后,Sohn等(1994)[6]报道了WSSMV法国分离物RNA1 3′-端4kb序列,于嘉林等(1995)[7]报道了一个中国河南分离物RNA1 3′-末端的891个核苷酸序列,发现与法国分离物仅有69.9%的序列同源性。最近,Namba等(1998)[8]报道了日本WYMV基因组全长序列,与法国分离物相比,也仅有69.8%的同源性,从而表明它们是不同的病毒。以下阐述这两种病毒的生物学和分子生物学特性。
, http://www.100md.com
1 生物学和生态学
1.1 宿主范围和症状
两种病毒在自然界都仅侵染普通小麦(Triticum aestivum)和硬粒小麦(T.durum)。曾有报道说WYMV在德国的黑麦(Secale cereale)上也有发现[9],然而,最近的研究表明该分离物是WSSMV[10]。在实验室条件下,两种病毒均能摩擦接种小麦和黑麦。
WYMV侵染的小麦在2月中旬显症,嫩叶上呈现褪绿条纹或黄花叶症状,在老叶上也常出现坏死斑。3~5月间气温升高后,花叶症状逐渐消失,新叶无症,但是植株分蘖减少。通常感病植株麦穗短小,发育不全。WSSMV症状通常为短褪绿或坏死斑点及梭条状条纹,引起小麦分蘖减少及矮化。温度是该类病害发展及症状表现的主要决定因素。
症状表现由于不同小麦品种而有所差别。通常感病品种症状严重,而抗病品种则无症或症状轻微。两种病害的田间症状都是田块呈现浅绿色到黄色,很容易和肥力缺乏引起的作物田间症状相混淆。
, 百拇医药
1.2 地理分布
土传线状小麦花叶病害在亚洲、欧洲和北美洲普遍发生,主要分布于北半球的冬小麦栽种区。已证实其存在的地区为:中国的安徽、河南、江苏、湖北、陕西、四川、山东和浙江等省,印度,日本的北海道、本州、九州、琉球和四国,加拿大的安大略省,美国的阿拉巴马、阿肯萨斯、乔治亚、伊利诺斯、印地安那、肯萨斯、肯特基、马里兰、密歇根、内布拉斯加、纽约、俄克拉哈马、宾西法尼亚和弗吉尼亚等州,意大利,法国和德国等。
通过不同地区分离物的基因组研究证实,欧洲和北美洲分离物是一个基因组同源的种,彼此间非常相似(93.5%~99.5%);中国和日本分离物是另一个基因组同源的种群(96.9%~99.0%);与欧美分离物具有很大的差异(核苷酸序列同源性67.7%~77.7%)。因此北美和欧洲分离物应当统一为WSSMV,而亚洲分离物应当为WYMV[10]。
1.3 病害传播
, http://www.100md.com
1.3.1 介体
禾谷多粘菌为根肿菌纲成员。Ledingham(1939)在加拿大安略省的小麦上首次发现该菌,通过9年观察,他对禾谷多粘菌的形态学和生活史作了详细的描述[11]。陈剑平等(1992,1993,1998)[12~14]和Littlefield等(1997)[15]对禾谷多粘菌超微结构作了详尽的研究。Castlebury和Domier(1998)[16]研究了根肿菌纲代表成员甘蓝根肿菌(Plasmodiophora brassicae)的进化史,他们在分析了核糖体小亚单位RNA基因(SSU rDNA)的序列数据后发现,它和对照的生物亲缘性都不高,但和原生动物的亲缘性要比它和真菌或是粘菌的亲缘性更高。禾谷多粘菌的宿主范围很广,包括禾谷类的大麦、小麦和燕麦,一些分离物还能侵染双子叶植物。
1.3.2 病毒-介体关系
, 百拇医药
Slykhuis和Barr(1978)[17]提出WSSMV是由禾谷多粘菌传播。他们用禾谷多粘菌接种小麦植株的根系,和(或)在植物叶片上摩擦接种病毒,然后将一些无病幼苗和它们栽种在一起,结果表明只有在WSSMV和禾谷多粘菌都存在的情况下,幼苗上才会发生病害,且该病害不能由其他测试的土栖真菌孢子传播。土壤中WSSMV的侵染性经52.5℃加热30min丧失,湿土或干土的侵染性在温室内或户外都可保持5年。
对于WYMV病害传播的研究不多。Ikata和Kawai(1940)[18]调查了WYMV病害的生态学,揭示该病害的病原是通过土壤传播的。带毒的土壤在户外放置5年或室内4年后丧失侵染性。因为WYMV和大麦黄花叶病毒(barley yellow mosaic virus,BaYMV)及WSSMV非常相似,故一般认为病毒存在于禾谷多粘菌体内以持续性方式传播。
1.3.3 病害流行学
, 百拇医药
秋季适当降雨和较低温度(4~13℃,最适为10℃)、冬季寒冷以及春季延续的低温有利于病害的发生和症状表现。连作感病小麦品种会导致该类病害的流行,并且病症加重,休耕能消除或降低土壤的侵染性。温度是病害发展及症状表现的主要决定因素[18,19]。
WSSMV及WYMV侵染冬小麦的最适温度为15℃(变动范围可在5~17℃,但不能超过20℃)。在适宜的温度及充足的土壤湿度条件下,小麦播种后4~10d病毒侵染就会发生,10~20d侵染达到高峰[18~20]。在北温带小麦栽种区,大约是在播种后10~40d,田间的病毒就随着禾谷多粘菌的游动孢子开始侵染小麦。播种30d后,用ELISA方法可以在冬小麦根部检测到病毒,60d后,就能在嫩叶上检测到病毒。
病毒侵染的植株数以及根组织中的病毒含量在秋冬季增加,嫩叶中的病毒含量在秋末、冬季和早春增加。通常这段时间的气温都低于5℃,表明病毒的二次侵染、复制、细胞-细胞间的运动以及系统性的运动都只在很低的温度条件下发生[20~22]。
, 百拇医药
这类病毒从根到叶的运动相当缓慢,约需30~40d,而从叶到根的运动仅需要5d。病毒从根到叶的运动之所以缓慢,可能取决于韧皮部对病毒的吸收情况[23]。
虽然秋冬季时ELISA法已能检测到植株中病毒的存在,但此时小麦植株是无症的[20~22]。冬末早春时分,温度常变动在5~13℃,植株经春化后重新生长,新叶才逐渐表现出症状。病土接种的植株,在合适温度下,通常需60d或更长的潜伏期才能表现出症状;即便是用病毒摩擦接种幼苗,症状表现也需要经过30~40d时间[24,25]。
用以下一些处理方法可以使潜伏期缩短几周,并使表现症状植株的百分数增加:1)交替的冰冻和融冻;2)培养温度在5~17℃范围内变动;3)严重冰冻(-17℃)损害后再生长;4)剪除植株嫩叶后再生长。除方法2外,其余方法都是对根(直接)和(或)叶片(直接或间接)的损害,推测可能是由于暂时性地改变了植株韧皮部结构,病毒更容易进入,从而导致潜伏期的缩短[23]。
, http://www.100md.com
1.3.4 经济影响
WSSMV和WYMV的危害主要表现为小麦分蘖减少、生长受阻、产量减少和质量下降。
在加拿大安大略省,WSSMV侵染引起的冬小麦产量损失一般在3%~59%之间。在美国北部、中部和东北部地区,WSSMV的侵染常导致24%~64%的减产。欧洲WSSMV通常是和土传小麦花叶病毒(soil-borne wheat mosaic virus,SBWMV)混合侵染小麦的,引起的作物产量损失也很严重。中国WYMV病害分布区域包括北部冬小麦区的胶东沿海区,黄淮冬小麦区的大部,长江中、下游冬小麦区的部分和西南冬小麦区的四川盆地区等小麦生态区。据我们1998年春天沿312国道从南京到河南潢川的调查,大约有20%~30%的小麦发病。如河南潢川县,WYMV常年发病面积在10000hm2;1991年大发生,发病面积23333hm2,3333hm2绝收;1970~1998年累计发病面积达206666hm2。一般病田小麦减产10%~30%,重病田减产可高达50%~70%。山东胶东半岛的小麦花叶病害是WYMV和中国小麦花叶病毒(Chinese wheat mosaic virus,CWMV)混合侵染引起的,危害更为严重,感病品种常常整片坏死[26]。在日本,WYMV可引起小麦产量的严重损失,高感病和轻微感病品种间存在35%的产量差异。
, http://www.100md.com
1.3.5 病害防治
由于这类病毒传播介体的休眠孢子抗逆性很强,化学试剂和杀菌剂的使用在经济和生态学上都是不可行的,休耕或轮作可以起到一定的防治作用,但是这类病害的控制主要靠栽种抗病品种。由于目前无法筛选到抗禾谷多粘菌的小麦品种,所以今后的育种仍有赖于病毒基因介导的抗性。浙江省农科院病毒室和德国马普所、中国农科院植保所及英国洛桑试验站合作,应用基因工程改良小麦,使小麦对这类病害产生抗性。目前全长病毒外壳蛋白和截短的NIb基因小麦表达载体已经构建完成,小麦转基因工作正在进行之中。
2 分子生物学
2.1 病毒粒子性质
WYMV和WSSMV均为弯曲的线状病毒粒子。WSSMV粒子典型长度为275~300nm和600~625nm,直径为12.8nm。提纯的线状病毒长度常常超过2000nm,可能是由于病毒粒子两端线性化聚集引起[4]。WYMV粒子长度典型分布峰为275~300nm和575~600nm,直径13~14nm[2]。
, 百拇医药
两种病毒都能在宿主植物中产生细胞质内含体。在WYMV侵染的小麦植株叶片细胞中已发现有复合膜状体、风轮体和圆柱状体的存在。这些内含体通常成簇出现并在原生质体中占据了很大的空间。病毒状粒子散布在原生质体中,或与小的结晶体及风轮状内含体结合[2,27]。WSSMV侵染的小麦细胞病理学和WYMV相似,膜状体通常是有序排列的。细胞质的某些部位存在很多风轮状内含体,与WYMV侵染细胞中的情况很相似。病毒状粒子散布或聚集在细胞质中,或结合有一些小的结晶体。粒子也沿着风轮状内含体的长轴方向并行排列[27,28]。
WYMV、WSSMV和BaYMV血清学相关。免疫电镜研究表明,BaYMV和WYMV抗血清反应较强,与美国WSSMV抗血清反应较弱,而与大麦和性花叶病毒(barley mild mosaic virus,BaMMV)或SBWMV抗血清不反应[10]。
2.2 基因组结构及编码的蛋白
, 百拇医药
两种病毒均具有两条单股、正链RNA基因组。WSSMV RNAs的分子量分别为2.6×106Da和1.4×106Da;WYMV RNAs的分子量分别为2.6×106Da和1.5×106Da。由WSSMV和WYMV外壳蛋白基因的核苷酸序列推测的外壳蛋白大小(CP)分别为31.9kDa和32kDa,和SDS-PAGE分析结果几乎相同[6,8,29]。WSSMV和WYMV的基因组结构相似,但同源性较低(图1和表1)。
图1 WSSMV-Fr和WYMV-T基因组结构图示及不同区域的同源性
Fig.1 Genomic organization and homology of WSSMV-Fr and WYMV-T
, 百拇医药
表1 WYMV-T和WSSMV-Fr的基因组结构及其编码蛋白
Table 1 Genomic organization and induced proteins of WYMV-T and WSSMV-Fr
WYMV-T
WSSMV-Fr
核苷酸
蛋白质
核苷酸
蛋白质
位置
大小
位置
, 百拇医药
大小
Mr(kD)
位置
大小
位置
大小
Mr(kD)
RNA1
5′-UTR
1~162
162
1~169
169
, http://www.100md.com
启始密
163~165
170~172
码子
P3
163~1143
981
1~327
327
36
170~1099
930
1~310
, http://www.100md.com
310
35
7kDa
1144~1341
198
328~393
66
7
1100~1297
198
311~376
66
7
, http://www.100md.com
CI
1342~3318
1977
394~1052
659
73
1298~3274
1977
377~1035
659
73
NIa
3319~4911
, 百拇医药
1593
1053~1583
531
60
3275~4970
1596
1036~1567
532
60
NIb
4912~6495
1584
1084~2111
, http://www.100md.com
528
59
4971~6454
1584
1568~2095
528
59
CP
6496~7374
879
2112~2404
293
32
, 百拇医药
6455~7336
882
2096~2389
294
32
ORF
163~7374
7212
1~2404
2404
269
170~7336
7167
, http://www.100md.com
1~2389
2389
267
终止子
7375~7377
7337~7339
3′-UTR
7378~7636
259
7340~7570
231
RNA2
, 百拇医药 168
5′-UTR
1~168
1~170
170
启始密
169~171
171~173
码子
P1
169~933
765
1~55
, 百拇医药
255
28
171~935
765
1~255
255
28
P2
934~2880
1947
256~904
649
73
, http://www.100md.com
936~2870
1935
256~900
635
71
ORF
169~2880
2712
1~904
904
101
171~2870
2700
, 百拇医药
1~900
900
100
终止子
2881~2883
2871~2873
3′-UTR
2884~3659
776
2874~4073
1200
WYMV-T(WYMV日本典型分离物)RNA1和RNA2分别由7636和3659nt组成[不包括3′-poly(A)尾]。计算机分析揭示,RNA1和RNA2分别具有一个长的开放阅读框架(ORF1和ORF2)。ORF1长7212nt,起始于nt 163~165处的第一个AUG,终止于nt 7375~7377处的UAA密码子。其后是长为259nt的3′-UTR。ORF1编码一个2404个氨基酸(aa)的蛋白质,分子量269kDa。推测269kDa蛋白经切割后产生6个蛋白,包括C端的CP和5个非结构蛋白。CP由293aa组成,分子量32kDa。NIb蛋白具有RNA依赖性的RNA聚合酶活性位点的保守模式SGX3TX3NT X18~37GDD(1915~1965aa)。NIa-Pro负责269kDa蛋白的切割。NIa-VPg含有一个Tyr,可能是起结合病毒RNA5′-末端的作用。CI蛋白具有NTP结合模式GXXGXGKS(486~583aa),不同处是第四个氨基酸G被V替换,被认为具有解螺旋酶的活性。P3蛋白具有锌指结构。7kDa蛋白功能仍不清楚。ORF2长为2712nt,起始于nt 169~171处的第一个AUG密码子,终止于nt 2881~2883处的UAG密码子,推测编码一个904aa、分子量101kDa的蛋白。101kDa蛋白在254~256aa处含有一个VGS三氨基酸切割位点,通过切割可产生两个大小分别为28kDa(P1)和73kDa(P2)的蛋白。P1蛋白含有蛋白酶结构域GYCY(141~144aa),并在215aa处有一个切割活性所必需的组氨酸残基,该蛋白和马铃薯Y病毒科其他成员的HC-Pro蛋白结构类似。P2蛋白含有一个假的CP结构域(319~398aa)和一个富含亮氨酸的结构域(595~777aa)。与马铃薯Y病毒科其他病毒的P2蛋白以及Furovirus的CP-readthrough(RT)蛋白具有一定的同源性。WYMV RNA1和RNA2的5′-UTR长度相似(分别为162和168nt),且具有相当的序列同源性(65%)。5′-UTR可能在病毒包装、复制及翻译中起着重要的作用。WYMV RNA1和2的3′-UTR在长度(分别为259和767nt)和序列上都具有很大差异[8,10]。
, http://www.100md.com
WSSMV-Fr(WSSMV法国分离物)RNA1和RNA2分别长为7570和4073核苷酸[不包括3′-poly(A)尾]。基因组结构和WYMV-T相似。RNA1 ORF1长7167核苷酸,起始于nt 170~172处的第一个AUG,终止于nt 7337~7339处的UAA密码子,编码一个2389aa的蛋白质,分子量266.5kDa;3′-UTR长231nt。ORF2长2700nt,起始于nt 171~173处的第一个AUG密码子,终止于nt 2871~2873处的UAG密码子,推测编码一个900aa、分子量99.8kDa的蛋白。两个聚合蛋白的结构和WYMV类似。RNA1和RNA2的5′-UTR长度相似(分别为169和170nt),3′-UTR在长度(分别为231和1200nt)和序列上都具有很大差异(表1)[6,30]。
2.3 分子进化和株系分化
WSSMV和WYMV外壳蛋白区域的序列同源性与确立的马铃薯Y病毒科不同种的划分标准是相符的,但较高的同源性也揭示WSSMV、WYMV和BaYMV是很相似的病毒。对于外壳蛋白区域核苷酸序列的相关性研究表明了这些病毒分离物间的近缘关系(图2)[10]。
, http://www.100md.com
1991年,在日本北海道地区Eniwa城发现一个有别于WYMV-T的株系,被命名为WYMV-H。在1991~1994年,Kusume等研究表明WYMV-T、WYMV-H和WSSMV在不同小麦品种上的致病性存在明显差异。WYMV-T能侵染关东地区的主要秋播小麦品种Norin 61,而WYMV-H不能;北海道地区的两种主要秋播小麦品种Chihokukomugi和Horoshirilomugi仅被WYMV-H侵染;且WYMV-H不侵染对WYMV-T高感病的品种Hatakedakomugi。ELISA结果也进一步表明了WYMV-T、WYMV-H和WSSMV之间的血清学差异,WYMV-H株系与WYMV-T在血清学上关系更近[3]。
图2 使用GCG程序PILEUP、DISTANCES和GROWTREE对WYMV、WSSMV和BaYMV各分离物的外壳蛋白核苷酸序列的分析,水稻坏死花叶病毒(rice necrosis mosaic virus,RNMV)作为一个根植的外围集团用TREEVIEW程序表示各分离物间的相关性。比例尺代表每100个碱基中不同碱基的数目[10]。
, http://www.100md.com
Fig.2 Phylogram of the coat protein nucleic acid sequences of isolates of WYMV,WSSMV and BaYMV prepared from the GCG programs PILEUP,DISTANCES and GROWTREE and displayed in TREEVIEW with RNMV as the rooted outgroup.The scale bar shows the number of substitutions per 100 bases.
我国WYMV分离物的部分序列测定表明它们的同源性较高,但初步田间试验表明不同试点的品种反应存在差异,因而可能存在不同于日本分离物的株系,有待进一步证实。
对于WSSMV的株系分化情况,至今尚无报道。
基金项目:863资助项目(101-04-01-05);浙江省重大科技项目(961106129);浙江省青年科技人才专项资金(RC9604)
, 百拇医药
作者简介:陈炯(1975年-),男,汉,浙江绍兴人,硕士,1999年入学读博士学位。主要从事植物病毒学研究。
参考文献
[1]Sawada E.Wheat yellow mosaic prevention[J].Journal of Plant Protection (Byochugai-Zasshi),1927,14:444~449
[2]Inouye T.Viral pathogen of the wheat yellow mosaic disease[J].Nogaku Kenkyu,1969,53:61~68
[3]Slykhuis JT.Evidence of soil-borne mosaic of wheat in Ontario[J].Canadian Plant Disease Survey,1960,40:43
, 百拇医药
[4]Slykhuis JT.Verification of wheat spindle streak mosaic virus in Ontario[J].Canadian Plant Disease Survey,1969,49:108~111
[5]Usugi T,Saito Y.Relationship between wheat yellow mosaic virus and wheat spindle streak mosaic virus[J].Annals of the Phytopathological Society of Japan,1979,45:397~400
[6]Sohn A,Schenk P,Signoret PA,et al.Sequence analysis of the 3′ terminal half of RNA1 of wheat spindle streak mosaic virus[J].Archives of Virology,1994,135:279~292
, http://www.100md.com
[7]于嘉林,晏立英,冯继东,等.一种中国发生的真菌传小麦花叶病毒RNA-1 3′末端核苷酸序列分析[J].病毒学报.1995,11(5):248~254
[8]Namba S,Kashiwazaki S,Lu X,et al.Complete nucleotide sequence of wheat yellow mosaic bymovirus genomic RNAs[J].Archives of Virology,1998,143,631~643
[9]Huth W,Lesemann D-E.Fungus-transmitted soil-borne viruses on rye in Germany[J].Journal of Plant Diseases and Protection,1993,103:120~124
[10]Jiong Chen,Anke Sohn,Jianping Chen,et al.Molecular comparisons amongst wheat bymovirus isolates from Asia,North America and Europe[J].(in submission)
, 百拇医药
[11]Leddingham GA.Studies on Polymyxa graminis,n.gen.n.sp.,a plasmodiophoraceous root parasite of wheat[J].Canadian Journal of Research C,1939,17:38~51
[12]陈剑平,Adams M J.禾谷多粘菌原质因和游动孢子囊的超微结构[J].浙江农业学报,1993,5:71~74
[13]陈剑平,Adams M J.禾谷多粘菌游动孢子超微结构[J].真菌学报,1992,11(3):216~220
[14]Chen JP,Wang Z,Hong J,et al.Ultrastructural studies of resting spore development in Polymyxa graminis[J].Mycol Res,1998,102(6):687-691
, 百拇医药
[15]Littlefield LJ,Delfosse P,Whallon JH,et al.Anatomy of sporosori of Polymyxa graminis,the vector of Indian peanut clump virus,in roots of Sorghum bicolor[J].Canadian Journal of Plant Pathology,1997,19:281~288
[16]Castlebury LA,Domier LL.Small subunit ribosomal RNA gene phylogeny of Plasmodiophora brassicae[J].Mycologia,1998,90:102~107
[17]Slykhuis JT,Barr DJS.Confirmation of Polymyxa graminis as a vector of wheat spindle streak mosaic virus[J].Phytopathology,1978,68:639~643
, http://www.100md.com
[18]Ikata S,Kawai I.Studies on wheat yellow mosaic disease[J].Noji-kairyo-shiryo,Ministry of Agriculture and Forestry,1940,154:1~123
[19]Slykhuis JT.Factors determining the development of wheat spindle streak mosaic caused by a soil-borne virus in Ontario[J].Phytopathology,1970,60:319~331
[20]Ohto Y,Naito S.Propagation of wheat yellow mosaic virus in winter wheat under low temperature conditions[J].Annals of the Phytopathological Society of Japan,1997,63:361~365
, http://www.100md.com
[21]Cunfer BM,Demski JW,Bays DC.Reduction in plant development,yield,and grainquality associated with wheat spindle streak mosaic virus[J].Phytopathology,1988,78:19~204
[22]Carroll JE[J],Bergstrom GC,Gray SM.Dynamics of wheat spindle streak mosaic bymovirus in winter wheat[J].European Journal of Plant Pathology,1997,103:313~321
[23]Schenck PM,Antoniw JF,Batista MdeF,et al.Movement of barley mild mosaic and baley yellow mosaic virus in leaves and roots of barley[J].Annals of Applied Biology,1995a,126:291~305
, http://www.100md.com
[24]Slykhuis JT.Factors critical to mechanical transmissibility of wheat spindle streak mosaic virus[J].Phytopathology,1975a,65:582~584
[25]Usugi T.Epidemiology and management in Japan of soil-borne cereal mosaic viruses with filamentous particles.In:Cooper JI,Asher MJC eds.Developments in Applied Biology 2,1988,Viruses with Fungal Vectors[M].Wellesbourne,UK:Association of Applied Biologists,1988.213~225
[26]Rong Ye,Tao Zheng,Jianping Chen,et al.Characterization and partial sequence of a new Furovirus of wheat in China[J].Plant Pathology,(Accepted)
, 百拇医药
[27]Hibino H,Usugi T,Saito Y.Comparative electron microscopy of inclusions associated with five soil-borne filamentous viruses of cereals[J].Ann Phytopath Soc Japan,1981,47:510~519
[28]Hooper GR,Wiese MV.Cytoplasmic inclusions in wheat affected by wheat spindle streak mosaic[J].Virology,1972,47:664~672
[29]Usugi T,Kashiwazaki S,Omura T,et al.Some properties of nucleic acids and coat proteins of soil-borne filamentous viruses[J].Ann Phytopath Soc Japan,1989,55:26~31
, 百拇医药
[30]Sohn A,Leclair S,Steinbiss H-H.Analysis of RNA1 of wheat spindle streak mosaic bymovirus (WSSMV)[C].In:Sherwood JL,Rush CM eds.Proceedings of the Third Symposium of the Internationsal Working Group on Plant Viruses with Fungal Vectors,Dundee.American Society of Sugaar Beet Technologists.USA:Denver,Co.1996.45~48
[31]Kusume T,Tamada T,Hattor H,et al.Identification of a new wheat yellow mosaic virus strain with specific pathogenicity towards major wheat cultivars grown in Hokkaido[J].Ann Phytopath Soc Japan,1997,63:107~109
收稿日期:1999-01-25
修回日期:1999-05-05, http://www.100md.com