L-�������Ԥ�����Է�Ѫ�ܽṹ�ع����Ƶ��о�
���ߣ��뽨�� �ž��� �Ա�
��λ���뽨�� �ž���(����ҽ�ƴ�ѧ��һҽԺ���� 100034)���Ա�(������ˮ̶ҽԺ�ڿ�)
�ؼ��ʣ�������ζ���������
�л�ҽѧ��־000325 ժ Ҫ��Ŀ�� ̽��L-������(L-Arg)��Ԥ�����Է�Ѫ�ܽṹ�ع��Ļ��ơ����� ��18ֻWistar������������������Ϊ�����顢������͵���+L-Arg��(��6��������)�������ĵ��ܷ��ⶨ�ζ���ѹ�������Դ������֯�걾�������ṹ�۲�ͳ��ṹ�۲죬ͬʱ�Էֹ��ȷ���ӲⶨѪ��һ������(NO)���������Է���֯����Ƥ��-1(ET-1) cRNA̽�����ԭλ�ӽ����о��ζ�����Ƥϸ��ET-1 mRNA�ı����� ���������ζ���ƽ��ѹ(mPAP)Ϊ2.71 kPa��0.29 kPa��������Ϊ2.05 kPa��0.14 kPa�����߱Ƚ�P<0.05������������Ѫ���������ṹ�������Ըı䣬�з�Ѫ�ܽṹ�ع��γɡ�ͬʱ���������Ѫ��NO��Ӻ���Ϊ3.54 ��mol/L��0.47 ��mol/L��������Ϊ4.79 ��mol/L��0.17 ��mol/L�����߱Ƚ�P<0.05��������ζ�����Ƥϸ��ET-1 mRNA����������ǿ��Ȼ��������+L-Arg�����mPAPΪ2.23 kPa��0.18 kPa��������Ϊ2.71 kPa��0.29 kPa�����߱Ƚ�P<0.05��L-Arg�����˵����Է�Ѫ�ܽṹ�ع����γɡ�ͬʱ����+L-Arg�����Ѫ��NO��Ӻ������Ը��ڵ�����(P<0.05)��L-Argʹ��������ζ�����Ƥϸ��ET-1 mRNA�������������ơ����� L-Argͨ���ٽ�������������NO���ɣ��Ӷ����Ʒζ�����Ƥϸ��ET-1 mRNA����Ե����Է�Ѫ�ܽṹ�ع��Լ������Էζ�����ѹ���γ�����Ҫ�ĵ������á�
, ��Ĵҽҩ
The mechanism responsible for alleviation of hypoxic pulmonary vascular structural remodeling by L-arginine
QI Jianguang DU Junbao ZHAO Bin.
(Department of Pediatrics, First Hospital, Beijing Medical University, Beijing 100034, China)
Abstract��Objective To explore the mechanism of the therapeutic effect of L-arginine on hypoxic pulmonary vascular structural remodeling. Methods Eighteen age- and body weight-matched Wistar rats were randomly divided into hypoxic group, hypoxic with L-arginine group or control group. Pulmonary artery mean pressure ( mPAP ) of each rat was evaluated using right cardiac catheterization. Pulmonary vascular microstructure was measured and the ultrastructural changes in intra-acinar pulmonary muscularized arteries were observed. Meanwhile, indirect plasma concentration of nitric oxide ( NO ) was measured via spectrophotometry, and endothelin-1(ET-1) mRNA expression in pulmonary artery endothelial cells was detected using in situ hybridization with a cRNA probe for ET-1. Results mPAP was significantly increased in hypoxic rats (2.71 kPa��0.29 kPa) as compared with that of normal controls (2.05 kPa��0.14 kPa)(P<0.05). Microstructure and ultrastructure of pulmonary arteries changed obviously in hypoxic rats with the development of hypoxic pulmonary vascular structural remodeling. Meanwhile, indirect plasma NO concentration in hypoxic rats (3.54 ��mol/L��0.47 ��mol/L ) was markedly decreased compared with controls (4.79 ��mol/L��0.17 ��mol/L)(P<0.05). The expression of ET-1 mRNA of hypoxic rats strengthened obviously. However, mPAP was significantly decreased in hypoxic rats treated with L-arginine (2.23 kPa��0.18 kPa ) as compared with that of hypoxic rats (2.71 kPa��0.29 kPa) (P<0.05). L-arginine ameliorated pulmonary vascular structural remodeling of hypoxic rats in association with an increase in indirect plasma NO concentration (P<0.05)and an inhibited ET-1 mRNA expression. Conclusion L-arginine plays an important role in the regulation of development of hypoxic pulmonary vascular remodeling and hypoxic pulmonary hypertension, promoting NO production and inhibiting ET-1 mRNA expression in hypoxic rats.
, ��Ĵҽҩ
Key words��Arginine; Pulmonary artery; Anoxia
�����Է�Ѫ�ܽṹ�ع����ٴ��ڶ��ķ�Ѫ�ܼ����ķ�����չ�м�Ϊ���еĹ�ϵ[1��2]��L-������(L-Arg)��(��)һ������(NO)���Ի�������Է�Ѫ�ܽṹ�ع��Լ������Էζ�����ѹ���γ�[3-5]�����������δ��ȫ���[6-8]�����о�ּ��̽��L-Arg��Ԥ�����Է�Ѫ�ܽṹ�ع������û��ƣ�Ϊ�ٴ���һ��Ӧ��L-Arg���ε����Էζ�����ѹ�ṩʵ�������
�����뷽��
1.������飺����Wistar����18ֻ������210��300 g������������������ѱ���ͬ�����ؽӽ���ÿ3ֻ�������1�������飬����ÿ��������Ĵ��������Ϊ�����顢������͵���+L-Arg�飬ÿ��6ֻ���������鼰����+L-Arg��������ڳ�ѹ������(�й�ҽѧ��ѧԺ����ҽѧ�о����ṩ)�ڣ����Ʋ�����Ũ����10.0%��0.5%��ÿ����������6 h�����������ܡ�����+L-Arg�����ÿ�����ǰ���̸�ǻ��ע��L-Arg 500 mg/kg�������鼰���������ÿ�츹ǻ��ע����ͬ�ݻ���������ˮ��3�����ij�����������ʳ������ͬ��
, ��Ĵҽҩ
2.����ζ���ѹ���IJⶨ�������������ܺ�ǻע��10%����̹(1 ml/100 mg)�������̶�������̨�ϣ��������ĵ��ܷ��ⶨ����ζ���ѹ����������ϩ���ܴ��Ҿ��⾲�����������Ҽ��ζ��������ܵ���һ�˾�P50ѹ����������RM-6000�Ͷർ��������������Ƿζ���ѹ�����ߣ��ⶨ�ζ���ƽ��ѹ(mPAP)��
3.����֯�걾��ȡ�ĺ��Ʊ����ⶨ�ζ���ѹ������ǻ��ȡ����һ���Ҷ��Һ���䶳��-70 �汣�档�ӱ�������֯�걾���ܲ�λ��ȡ����1 mm��1 mm��1 mm����֯�飬�����ȩ���������˫�̶���ϵ�б�ͪ��ˮ��Epon812���뱡��Ƭ��붨λ��ѡȡ�⾶Ϊ15��150 ��m�ķ��С�С���Ͷ����г�����Ƭ������˫����-������Ǧ˫Ⱦɫ��100CX-II����������۲졣��һ���Ҷ��10%��ȩ����Һ�̶����ݶȾƾ�˳����ˮ�����ױ�����ʯ��������Ƭ(��5 ��m)��������Ƭ��Weight������άȾɫ��Van Gieson��Ⱦ����������Ƭ����ԭλ�ӽ���
4.���ͷ�СѪ�ܰٷֱȵIJⶨ����40���⾵�£��۲첢����������Ƭ��СѪ��(�⾶>15 ��m����50 ��m)�м��Ͷ��������ּ��Ͷ������Ǽ���Ѫ�ܵ���Ŀ���ֱ�������Ǹ�ռ��СѪ�������İٷֱȡ����Ͷ����ڹ⾵�¾����������������㵯���㣻���ּ��Ͷ����ڹ⾵�¾����������ⵯ���㣬���ڵ����㲻�������Ǽ���Ѫ���ڹ⾵�½���һ�㵯���㡣
, ��Ĵҽҩ
5.���С�С���Ͷ��������Ĥ��ȼ�����IJⶨ��Ӧ��Q550 LWͼ���������ϵͳ��������Ƭ���ڵ�������������״�Ϲ���ķ��С�С���Ͷ������з���������Leica Qwin��������ÿ��Ѫ�ܵ�����ָ�꣺����Ѫ�����ĵ��ⵯ����֮�����⾶������⾶���ڵ����㳤�ȣ��ڵ������Χ��������ⵯ�����Χ�����������Barth��[9]�ķ������㲻ͬ����Ƕȼ����ڲ�ͬ����״̬��Ѫ��֮�����Ĥ���(RMT)�������Ĥ���(RMA)��ÿֻ�������5��10�����С�С���Ͷ������ֱ������еȼ��Ͷ���(�⾶>50 ��m����150 ��m)�ͷ�С���Ͷ���(�⾶>15 ��m����50 ��m)RMT��RMA���������ֵ��
6.Ѫ��NO��Ӻ����ⶨ��������ǰ���Ծ��ܶ���ȡѪ2 ml�����ĺ���ȡѪ��0.1 ml����9.3 mmol/L����п0.6 ml��˫��ˮ0.4 ml������1 mol/L��������0.1 ml����Ϸ���1 h�����ĺ�ȡ����Һ0.6 ml����˫��ˮ��3 g/L�ǰ�����6 mmol/L����ϩ���������η�Ӧ1 h����545 nm��ɫ�������ܶ�ֵ������������NO��Ӻ�����
, ��Ĵҽҩ
7.ԭλ�ӽ�������Ƥ��-1(ET-1) cDNA(����1.1 Kb)������ת¼�ķ�������������ر�ǵ�ET-1 cRNA̽�룬�Ը���������֯�걾��ԭλ�ӽ����걾�����ױ��������ݶ��Ҵ���������ˮ�������Ỻ����Һ(PBS)ϴ��1 min������������0.2 mol/L���ᴦ��10 min����PBSϴ��2 min��2�κ��Ե���øK(50 ��g/ml)��20 min��37�洦������40 g/L��ۼ�ȩ�������¹̶�10 min��ÿƬ�μ��ӽ�Һ25 ��l����ʯ�����Ĥ���ǣ���42�汣��24 h��ϵ�б���������-�Ȼ���Һ(SSC)ϴ�ӣ���1��100��Ѫ�嵰��ա����������ر��cRNA̽��ԭλ�ӽ���Ƭ�м�1��100 ABC����Һ������1 h���Թ�������ø-������(DAB)��ɫ��ϵ����ѧ�����¼��ԭλ�ӽ������źţ������ð붨������[10]����ET-1mRNA�������з������ػ�ɫ����Ϊ�ӽ������źš���һ�ζ�����Ƥϸ���������ź���Ϊ����(-)��1%��50%���������ź���Ϊ����(+)��51%��100%���������ź���Ϊǿ����(++)����ÿֻ�������֯��Ƭ�۲�15��20ֻ�ζ��������Զ���Ƭ�ӽ�ʱ����ET-1cRNA̽�룬��������������ȫ��ͬ��
, ��Ĵҽҩ
8.ͳ��ѧ�����������������Լ������˫���ط��������q���顣
���
1.�ζ���ѹ��(��1)��mPAP��3�������������������(P<0.05)�����������mPAP���Ը��ڶ�����(q=5.12, P<0.05)������+L-Arg�����mPAP���Ե��ڵ�����(q=3.72, P<0.05)���� ��������Ȳ�������������(q=1.41, P>0.05)��
��1 3�����ζ���ƽ��ѹ���ζ������ṹ��Ѫ��һ��������Ӻ����ı仯(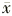 ��
��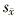 ) ���
) ���
����(ֻ)
mPAP
, ��Ĵҽҩ
(kPa)
���ͷ�СѪ�ܰٷֱ�(%)
�����Ĥ���(%)
�����Ĥ���(%)
Ѫ��NO���
����(��mol/L)
MA
PMA
NMV
���е�MA
��СMA
���е�MA
, http://www.100md.com
��СMA
������
6
2.05��
0.14
9.33��
0.53
7.07��
0.25
83.59��
0.56
7.70��
0.41
, ��Ĵҽҩ
10.12��
0.60
14.76��
0.75
19.18��
1.08
4.79��
0.17
������
6
2.71��
0.29*
16.04��
, http://www.100md.com
0.74**
10.32��
0.35**
73.64��
0.73**
9.94��
0.30**
15.15��
0.21**
18.87��
0.54**
, ��Ĵҽҩ
27.99��
0.36**
3.54��
0.47��
����+
L-Arg��
6
2.23��
0.18��
11.81��
0.24������
9.30��
, http://www.100md.com
0.18����
78.50��
0.25**��
8.28��
0.12��
11.86��
0.53����
15.84��
0.22��
22.26��
0.93*��
, ��Ĵҽҩ
5.21��
0.26��
Fֵ
6.90
54.25
33.35
80.39
15.69
39.57
15.88
38.05
7.65
Pֵ
, ��Ĵҽҩ
<0.05
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.01
ע��(1)mPAP���ζ���ƽ��ѹ��MA�����Ͷ�����PMA�����ּ��Ͷ�����NMV���Ǽ���Ѫ�ܣ�(2)�������Ƚ�*P<0.05��**P<0.01���������Ƚϣ���P<0.05����P<0.01��(3)Fֵ��PֵΪ���Ƚ�
, ��Ĵҽҩ
2.���ͷ�СѪ�ܰٷֱ�(��1)�����ͷ�СѪ�ܰٷֱ���3�����֮����������������(P��<0.001)������������СѪ�ܼ��Ͷ����ٷֱ����Ը��ڶ�����(q=14.57, P<0.01)������+L-Arg������СѪ�ܼ��Ͷ����ٷֱ����Ե��ڵ�����(q=9.19, P<0.01)�����Ը��ڶ�����(q=5.38, P<0.01)������������СѪ�ܷǼ���Ѫ�ܰٷֱ����Ե��ڶ�����(q=17.99,P<0.01), ����+L-Arg������СѪ�ܷǼ���Ѫ�ܰٷֱ����Ը��ڵ�����(q=8.74,P<0.01)�����Ե��ڶ�����(q=9.19, P<0.01)��
3.���С�С���Ͷ��������Ĥ��ȼ����(��1)�����С�С���Ͷ��������Ĥ��ȼ������3�����֮����������������(P��<0.001)�������������еȼ��Ͷ���RMT��RMA���Ը��ڶ�����(q=7.63��7.69, P��<0.01)����С���Ͷ���RMT��RMAҲ���Ը��ڶ�����(q=12.39,12.16, P��<0.01)������+L-Arg�������еȼ��Ͷ���RMT��RMA���Ե��ڵ�����(q=5.65, 5.65,P��<0.01)�������������Ȳ�������������(q=1.98��2.04, P��>0.05)����С���Ͷ���RMT��RMA���Ե��ڵ�����(q=8.09, 7.9, P��<0.01)�����Ը��ڶ�����(q=4.3, 4.25, P��<0.05)��
, ��Ĵҽҩ
4.���С�С���Ͷ�����Ƥϸ�����ṹ�������������С�С���Ͷ�����Ƥϸ�������ƽ���ڳ�Ѫ���ڱڣ��ṹ������ƽ����ϸ������С�����������ͣ�������������˿ռ�ݣ������������ܰ�(ͼ1)�����������ζ�����Ƥϸ���������ͣ�����ͻ���ǻ�����ı�ʽ������Ƥϸ�����ͣ������ڿɼ��������ݣ����������ͣ����������ţ�ƽ����ϸ������ʴ�ͬ�̶���ϳɱ���ת���������м�˿�����ܰ��٣�����������������������ϸ��������(ͼ2)������+L-Arg�����ζ�����Ƥϸ�������ƽ���ڳ�Ѫ���ڱڣ������ͳ̶Ƚϵ�������ᣬ�������Լ��٣�����ƽ����ϸ�������������ͣ��������Դ��ڹ��ɱ���(ͼ3)��
ͼ1 ���������μ��Ͷ�������Ƥϸ����ƽ���ṹ������ƽ����ϸ�����������ͣ�������������˿�����ܰ���ռ�� ��10 000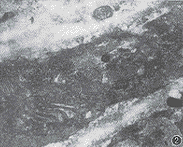
, http://www.100md.com
ͼ2 ���������μ��Ͷ�������Ƥϸ�����ͣ������ڼ��������ݡ�ƽ����ϸ���ʺϳɱ��ͣ������ڿɼ�������������壬������������� ��20 000
ͼ3 ����+L-Arg�����μ��Ͷ�������Ƥϸ����ƽ���������Լ��١�ƽ����ϸ������������ ��20 000
5.Ѫ��NO�ļ�Ӻ���(��1)��Ѫ��NO��Ӻ�����3�����֮���������������(P<0.001)�����������Ѫ��NO��Ӻ������Ե��ڶ�����(q=3.94, P<0.05)������+L-Arg�����Ѫ��NO��Ӻ������Ը��ڵ�����(q=5.22, P<0.05), ���������Ƚϲ�������������(q=1.33,P>0.05)��
6.�ζ�����Ƥϸ��ET-1 mRNA������������ַζ���(77%��7%)��Ƥϸ��ET-1 mRNA����������ź�(+)(ͼ4)����������(14%��7%)������+L-Arg��(4%��4%)����������ζ�����Ƥϸ����ʾET-1 mRNA���������ź�(+)��������зdz���������(q=9.32,10.86��P��<0.01)���������鼰����+L-Arg�����ζ�����Ƥϸ��ET-1 mRNA�����������������(q=1.54��P>0.05)��������(86%��7%)������+L-Arg��(96%��4%)����ַζ�����Ƥϸ����ET-1 mRNA����(ͼ5)�������������������ζ���(23%��7%)��Ƥϸ����ET-1 mRNA���������зdz���������(q=9.32, q=10.86��P��<0.01)���������鼰����+L-Arg�����ζ�����Ƥϸ��ET-1 mRNA�����������������(q =1.54��P>0.05)��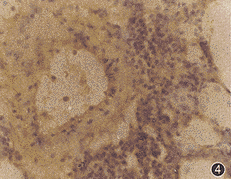
, ��Ĵҽҩ
ͼ4 ��������ζ�����1%��50%��Ƥϸ����ʾET-1 mRNA�ӽ������ź� DAB��ɫ����250
ͼ5 ����+L-Arg�����ζ�����Ƥϸ��ET-1 mRNA���������ܵ����� DAB��ɫ����400
����
�����Էζ�����ѹ���ٴ��ڶ��ķμ���������չ��������Ҫ�IJ�����������[1]�������Է�Ѫ�ܽṹ�ع������Ե����Էζ�����ѹ����Ҫ��������������Ҫ�����Ƿζ���ƽ����ϸ���������ʺ���Ĥ��������СѪ�ܼ������Լ�ϸ������ʵ�����[2]�����о����֣���ѹ����2�ܴ���ζ���ѹ���������ߣ���Ѫ�ܽṹ�ع����⾵�·�СѪ���м����̶�������ǿ�����С�С���Ͷ���RMT��RMA�������ӣ��羵�·ζ�����Ƥϸ���������ʺ����ͣ���Ĥƽ����ϸ���������ʴ���ͬ�̶ȵ���ϳɱ���ת����ͬʱ����2�ܺ����Ѫ��NO��Ӻ������Խ��ͣ���ET-1 mRNA����������ǿ����ʾ������NOˮƽ�Ľ��ͺ�ET-1����������ǿ���ܲ����˵����Է�Ѫ�ܽṹ�ع��������Էζ�����ѹ���γɡ��ɴ��������ܽ�NOˮƽ���ߺ�/��ET-1ˮƽ���Ϳ��Է��ε����Է�Ѫ�ܽṹ�ع���Ϊ�ٴ����Ƶ����Էζ�����ѹ�ṩ��һ��˼·��
, ��Ĵҽҩ
�о���������Դ�Ը���L-Arg��(��)NO���Ի�������Է�Ѫ�ܽṹ�ع��Լ������Էζ�����ѹ���γ�[3-5]���ٴ����ѿ�ʼӦ��NO����ķ��������Ƴ���̥��ѭ����̥�������ۺ����ȼ���������ȡ�ó�����Ч[11]����NO������øߣ������������ӣ�����һ���Ķ������ã�������������ٴ�Ӧ�á�
L-ArgΪNO�ϳɵ�ǰ�壬���������ں�NO��ø������NO���������Ӧ��L-Arg��Ԥ�����Է�Ѫ�ܽṹ�ع����о��õ�Խ��Խ������ӣ����������δ��ȫ���������ʵ��֤ʵ��NO��Ѫ��ƽ����ϸ������ֳ��ֱ����������[12]����ʾL-Arg��Ԥ�����Է�Ѫ�ܽṹ�ع��Ļ���֮һ������ͨ��NO��ֱ�����á���һ��������ʵ�黹��ʾ��NO�������꾲����Ƥϸ��ET-1�ĺϳ�����ڣ����ɽ���Ѫ��ET-1ˮƽ[6]�������������о����Ҳ��ʾ��NO �����Ե�������ζ���ET-1�����������������[7]����ET-1��Ŀǰ���ֵ�������ǿ����Ѫ�ܻ��Զ��ģ����Ҿ���ǿ�ҵĴ�Ѫ��ƽ������ֳ������[8]���ɴ˼����ʾL-Arg��Ԥ�����Է�Ѫ�ܽṹ�ع�����һ���ܻ�����ͨ���ٽ�NO�ϳɣ��Ӷ��Դٽ�Ѫ��ƽ����ϸ����ֳ����ET-1���������á�
, ��Ĵҽҩ
Ϊ��ȷ��һ���ڻ��ƵĿ����ԣ����ǽ�L-Arg�����Ե����Էζ�����ѹ�����Ѫ�����ͳ��ṹ��Ѫ��NO�ļ�Ӻ����Լ��ζ�����Ƥϸ��ET-1 mRNA�����Ӱ�������ͬ���о������������L-Argʹ�����Էζ�����ѹ����ζ���ѹ�����Խ��ͣ���СѪ�ܼ����̶����Լ��ᣬ���С�С���Ͷ���RMT��RMA���Խ��ͣ��ζ������ṹ�ĸı����Ի��⣻ͬʱL-Argʹ��������Ѫ��NO��Ӻ����������߲���ʹ��������ζ�����Ƥϸ��ET-1 mRMA�������������ơ���ʾL-Argͨ���ٽ�����NO���ɣ��Ӷ����Ʒζ�����Ƥϸ��ET-1 mRMA����Ե����Է�Ѫ�ܽṹ�ع��Լ������Էζ�����ѹ���γ�����Ҫ�ĵ������á�
������Ŀ��������Ȼ��ѧ����������Ŀ(39470735, 39870844)
�ο����ף�
1 �ž���, ����. ȱ���Էζ�����ѹ. ����������ҽ�ƴ�ѧ���й�Э��ҽ�ƴ�ѧ���ϳ�����, 1994. 79.
, http://www.100md.com
2 Vender RL. Chronic hypoxic pulmonary hypertension: cell biology to pathophysiology. Chest, 1994, 106: 236-243.
3 �뽨�⣬�ž���. ��Դ��һ���������ڴ�������Է�Ѫ�ܽṹ�ع�. �л�������־��1999��37��104-106.
4 �ֺ�, ��Ӣ��, ��Ϊ, ��. L-������������ƶ�ȱ������ζ���ѹ��Ѫ�ܽṹ�仯��Ӱ�� . �й�ҽѧ��ѧԺѧ�� , 1997 , 19 : 179-184.
5 Mitani Y, Maruyama K, Sakurai M. Prolonged administration of L-Arginine ameliorates chronic pulmonary hypertension and pulmonary vascular remodeling in rats. Circulation, 1997, 96: 689-697.
, ��Ĵҽҩ
6 Kourembanas S, Mcquillan LP, Leung GK. et al. Nitric oxide regulates the expression of vasoconstrictors and growth factors by vascular endothelium under both normoxia and hypoxia. J Clin Invest, 1993, 92: 99-104.
7 �ž���, ������, �Ա�, ��. һ������������ȱ������ζ�����Ƥ�ػ�������Ӱ�� . �л�ҽѧ��־ , 1997 , 77 : 263-265.
8 Janakidevi K, Fisher MA, Del Vecchio PJ, et al. Endothelin-1 stimulates DNA synthesis and proliferation of pulmonary artery smooth muscle cells. Am J Physiol, 1992, 263: C1295-C1301.
, http://www.100md.com
9 Barth PJ, Kimpel C, Roy S, et al. An improved mathematical approach for the assessment of the medial thickness of pulmonary arteries. Path Res Pract, 1993, 189: 567-576.
10 Hislop AA, Springall DR, Buttery LDK, et al. Abundance of endothelial nitric oxide synthase in newborn intrapulmonary arteries. Arch Dis Child, 1995, 73: F17-F21.
11 Roberts JD, Polaner DM, Lang P, et al. Inhaled nitric oxide reverses hypoxemia in persistent pulmonary hypertension of the newborns. Lancet, 1992, 340: 818-819.
12 Garg UC, Hassid A. Nitric oxide-generating vasodilators and 8-bromo-cyclic guanosine monophosphate inhibit mitogenesis and proferation of cultured rat vascular smooth muscle cells. J Clin Invest, 1989, 83: 1774-1777.
�ո����ڣ�1999-08-24, http://www.100md.com
��λ���뽨�� �ž���(����ҽ�ƴ�ѧ��һҽԺ���� 100034)���Ա�(������ˮ̶ҽԺ�ڿ�)
�ؼ��ʣ�������ζ���������
�л�ҽѧ��־000325 ժ Ҫ��Ŀ�� ̽��L-������(L-Arg)��Ԥ�����Է�Ѫ�ܽṹ�ع��Ļ��ơ����� ��18ֻWistar������������������Ϊ�����顢������͵���+L-Arg��(��6��������)�������ĵ��ܷ��ⶨ�ζ���ѹ�������Դ������֯�걾�������ṹ�۲�ͳ��ṹ�۲죬ͬʱ�Էֹ��ȷ���ӲⶨѪ��һ������(NO)���������Է���֯����Ƥ��-1(ET-1) cRNA̽�����ԭλ�ӽ����о��ζ�����Ƥϸ��ET-1 mRNA�ı����� ���������ζ���ƽ��ѹ(mPAP)Ϊ2.71 kPa��0.29 kPa��������Ϊ2.05 kPa��0.14 kPa�����߱Ƚ�P<0.05������������Ѫ���������ṹ�������Ըı䣬�з�Ѫ�ܽṹ�ع��γɡ�ͬʱ���������Ѫ��NO��Ӻ���Ϊ3.54 ��mol/L��0.47 ��mol/L��������Ϊ4.79 ��mol/L��0.17 ��mol/L�����߱Ƚ�P<0.05��������ζ�����Ƥϸ��ET-1 mRNA����������ǿ��Ȼ��������+L-Arg�����mPAPΪ2.23 kPa��0.18 kPa��������Ϊ2.71 kPa��0.29 kPa�����߱Ƚ�P<0.05��L-Arg�����˵����Է�Ѫ�ܽṹ�ع����γɡ�ͬʱ����+L-Arg�����Ѫ��NO��Ӻ������Ը��ڵ�����(P<0.05)��L-Argʹ��������ζ�����Ƥϸ��ET-1 mRNA�������������ơ����� L-Argͨ���ٽ�������������NO���ɣ��Ӷ����Ʒζ�����Ƥϸ��ET-1 mRNA����Ե����Է�Ѫ�ܽṹ�ع��Լ������Էζ�����ѹ���γ�����Ҫ�ĵ������á�
, ��Ĵҽҩ
The mechanism responsible for alleviation of hypoxic pulmonary vascular structural remodeling by L-arginine
QI Jianguang DU Junbao ZHAO Bin.
(Department of Pediatrics, First Hospital, Beijing Medical University, Beijing 100034, China)
Abstract��Objective To explore the mechanism of the therapeutic effect of L-arginine on hypoxic pulmonary vascular structural remodeling. Methods Eighteen age- and body weight-matched Wistar rats were randomly divided into hypoxic group, hypoxic with L-arginine group or control group. Pulmonary artery mean pressure ( mPAP ) of each rat was evaluated using right cardiac catheterization. Pulmonary vascular microstructure was measured and the ultrastructural changes in intra-acinar pulmonary muscularized arteries were observed. Meanwhile, indirect plasma concentration of nitric oxide ( NO ) was measured via spectrophotometry, and endothelin-1(ET-1) mRNA expression in pulmonary artery endothelial cells was detected using in situ hybridization with a cRNA probe for ET-1. Results mPAP was significantly increased in hypoxic rats (2.71 kPa��0.29 kPa) as compared with that of normal controls (2.05 kPa��0.14 kPa)(P<0.05). Microstructure and ultrastructure of pulmonary arteries changed obviously in hypoxic rats with the development of hypoxic pulmonary vascular structural remodeling. Meanwhile, indirect plasma NO concentration in hypoxic rats (3.54 ��mol/L��0.47 ��mol/L ) was markedly decreased compared with controls (4.79 ��mol/L��0.17 ��mol/L)(P<0.05). The expression of ET-1 mRNA of hypoxic rats strengthened obviously. However, mPAP was significantly decreased in hypoxic rats treated with L-arginine (2.23 kPa��0.18 kPa ) as compared with that of hypoxic rats (2.71 kPa��0.29 kPa) (P<0.05). L-arginine ameliorated pulmonary vascular structural remodeling of hypoxic rats in association with an increase in indirect plasma NO concentration (P<0.05)and an inhibited ET-1 mRNA expression. Conclusion L-arginine plays an important role in the regulation of development of hypoxic pulmonary vascular remodeling and hypoxic pulmonary hypertension, promoting NO production and inhibiting ET-1 mRNA expression in hypoxic rats.
, ��Ĵҽҩ
Key words��Arginine; Pulmonary artery; Anoxia
�����Է�Ѫ�ܽṹ�ع����ٴ��ڶ��ķ�Ѫ�ܼ����ķ�����չ�м�Ϊ���еĹ�ϵ[1��2]��L-������(L-Arg)��(��)һ������(NO)���Ի�������Է�Ѫ�ܽṹ�ع��Լ������Էζ�����ѹ���γ�[3-5]�����������δ��ȫ���[6-8]�����о�ּ��̽��L-Arg��Ԥ�����Է�Ѫ�ܽṹ�ع������û��ƣ�Ϊ�ٴ���һ��Ӧ��L-Arg���ε����Էζ�����ѹ�ṩʵ�������
�����뷽��
1.������飺����Wistar����18ֻ������210��300 g������������������ѱ���ͬ�����ؽӽ���ÿ3ֻ�������1�������飬����ÿ��������Ĵ��������Ϊ�����顢������͵���+L-Arg�飬ÿ��6ֻ���������鼰����+L-Arg��������ڳ�ѹ������(�й�ҽѧ��ѧԺ����ҽѧ�о����ṩ)�ڣ����Ʋ�����Ũ����10.0%��0.5%��ÿ����������6 h�����������ܡ�����+L-Arg�����ÿ�����ǰ���̸�ǻ��ע��L-Arg 500 mg/kg�������鼰���������ÿ�츹ǻ��ע����ͬ�ݻ���������ˮ��3�����ij�����������ʳ������ͬ��
, ��Ĵҽҩ
2.����ζ���ѹ���IJⶨ�������������ܺ�ǻע��10%����̹(1 ml/100 mg)�������̶�������̨�ϣ��������ĵ��ܷ��ⶨ����ζ���ѹ����������ϩ���ܴ��Ҿ��⾲�����������Ҽ��ζ��������ܵ���һ�˾�P50ѹ����������RM-6000�Ͷർ��������������Ƿζ���ѹ�����ߣ��ⶨ�ζ���ƽ��ѹ(mPAP)��
3.����֯�걾��ȡ�ĺ��Ʊ����ⶨ�ζ���ѹ������ǻ��ȡ����һ���Ҷ��Һ���䶳��-70 �汣�档�ӱ�������֯�걾���ܲ�λ��ȡ����1 mm��1 mm��1 mm����֯�飬�����ȩ���������˫�̶���ϵ�б�ͪ��ˮ��Epon812���뱡��Ƭ��붨λ��ѡȡ�⾶Ϊ15��150 ��m�ķ��С�С���Ͷ����г�����Ƭ������˫����-������Ǧ˫Ⱦɫ��100CX-II����������۲졣��һ���Ҷ��10%��ȩ����Һ�̶����ݶȾƾ�˳����ˮ�����ױ�����ʯ��������Ƭ(��5 ��m)��������Ƭ��Weight������άȾɫ��Van Gieson��Ⱦ����������Ƭ����ԭλ�ӽ���
4.���ͷ�СѪ�ܰٷֱȵIJⶨ����40���⾵�£��۲첢����������Ƭ��СѪ��(�⾶>15 ��m����50 ��m)�м��Ͷ��������ּ��Ͷ������Ǽ���Ѫ�ܵ���Ŀ���ֱ�������Ǹ�ռ��СѪ�������İٷֱȡ����Ͷ����ڹ⾵�¾����������������㵯���㣻���ּ��Ͷ����ڹ⾵�¾����������ⵯ���㣬���ڵ����㲻�������Ǽ���Ѫ���ڹ⾵�½���һ�㵯���㡣
, ��Ĵҽҩ
5.���С�С���Ͷ��������Ĥ��ȼ�����IJⶨ��Ӧ��Q550 LWͼ���������ϵͳ��������Ƭ���ڵ�������������״�Ϲ���ķ��С�С���Ͷ������з���������Leica Qwin��������ÿ��Ѫ�ܵ�����ָ�꣺����Ѫ�����ĵ��ⵯ����֮�����⾶������⾶���ڵ����㳤�ȣ��ڵ������Χ��������ⵯ�����Χ�����������Barth��[9]�ķ������㲻ͬ����Ƕȼ����ڲ�ͬ����״̬��Ѫ��֮�����Ĥ���(RMT)�������Ĥ���(RMA)��ÿֻ�������5��10�����С�С���Ͷ������ֱ������еȼ��Ͷ���(�⾶>50 ��m����150 ��m)�ͷ�С���Ͷ���(�⾶>15 ��m����50 ��m)RMT��RMA���������ֵ��
6.Ѫ��NO��Ӻ����ⶨ��������ǰ���Ծ��ܶ���ȡѪ2 ml�����ĺ���ȡѪ��0.1 ml����9.3 mmol/L����п0.6 ml��˫��ˮ0.4 ml������1 mol/L��������0.1 ml����Ϸ���1 h�����ĺ�ȡ����Һ0.6 ml����˫��ˮ��3 g/L�ǰ�����6 mmol/L����ϩ���������η�Ӧ1 h����545 nm��ɫ�������ܶ�ֵ������������NO��Ӻ�����
, ��Ĵҽҩ
7.ԭλ�ӽ�������Ƥ��-1(ET-1) cDNA(����1.1 Kb)������ת¼�ķ�������������ر�ǵ�ET-1 cRNA̽�룬�Ը���������֯�걾��ԭλ�ӽ����걾�����ױ��������ݶ��Ҵ���������ˮ�������Ỻ����Һ(PBS)ϴ��1 min������������0.2 mol/L���ᴦ��10 min����PBSϴ��2 min��2�κ��Ե���øK(50 ��g/ml)��20 min��37�洦������40 g/L��ۼ�ȩ�������¹̶�10 min��ÿƬ�μ��ӽ�Һ25 ��l����ʯ�����Ĥ���ǣ���42�汣��24 h��ϵ�б���������-�Ȼ���Һ(SSC)ϴ�ӣ���1��100��Ѫ�嵰��ա����������ر��cRNA̽��ԭλ�ӽ���Ƭ�м�1��100 ABC����Һ������1 h���Թ�������ø-������(DAB)��ɫ��ϵ����ѧ�����¼��ԭλ�ӽ������źţ������ð붨������[10]����ET-1mRNA�������з������ػ�ɫ����Ϊ�ӽ������źš���һ�ζ�����Ƥϸ���������ź���Ϊ����(-)��1%��50%���������ź���Ϊ����(+)��51%��100%���������ź���Ϊǿ����(++)����ÿֻ�������֯��Ƭ�۲�15��20ֻ�ζ��������Զ���Ƭ�ӽ�ʱ����ET-1cRNA̽�룬��������������ȫ��ͬ��
, ��Ĵҽҩ
8.ͳ��ѧ�����������������Լ������˫���ط��������q���顣
���
1.�ζ���ѹ��(��1)��mPAP��3�������������������(P<0.05)�����������mPAP���Ը��ڶ�����(q=5.12, P<0.05)������+L-Arg�����mPAP���Ե��ڵ�����(q=3.72, P<0.05)���� ��������Ȳ�������������(q=1.41, P>0.05)��
��1 3�����ζ���ƽ��ѹ���ζ������ṹ��Ѫ��һ��������Ӻ����ı仯(
��) �������(ֻ)
mPAP
, ��Ĵҽҩ
(kPa)
���ͷ�СѪ�ܰٷֱ�(%)
�����Ĥ���(%)
�����Ĥ���(%)
Ѫ��NO���
����(��mol/L)
MA
PMA
NMV
���е�MA
��СMA
���е�MA
, http://www.100md.com
��СMA
������
6
2.05��
0.14
9.33��
0.53
7.07��
0.25
83.59��
0.56
7.70��
0.41
, ��Ĵҽҩ
10.12��
0.60
14.76��
0.75
19.18��
1.08
4.79��
0.17
������
6
2.71��
0.29*
16.04��
, http://www.100md.com
0.74**
10.32��
0.35**
73.64��
0.73**
9.94��
0.30**
15.15��
0.21**
18.87��
0.54**
, ��Ĵҽҩ
27.99��
0.36**
3.54��
0.47��
����+
L-Arg��
6
2.23��
0.18��
11.81��
0.24������
9.30��
, http://www.100md.com
0.18����
78.50��
0.25**��
8.28��
0.12��
11.86��
0.53����
15.84��
0.22��
22.26��
0.93*��
, ��Ĵҽҩ
5.21��
0.26��
Fֵ
6.90
54.25
33.35
80.39
15.69
39.57
15.88
38.05
7.65
Pֵ
, ��Ĵҽҩ
<0.05
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.01
ע��(1)mPAP���ζ���ƽ��ѹ��MA�����Ͷ�����PMA�����ּ��Ͷ�����NMV���Ǽ���Ѫ�ܣ�(2)�������Ƚ�*P<0.05��**P<0.01���������Ƚϣ���P<0.05����P<0.01��(3)Fֵ��PֵΪ���Ƚ�
, ��Ĵҽҩ
2.���ͷ�СѪ�ܰٷֱ�(��1)�����ͷ�СѪ�ܰٷֱ���3�����֮����������������(P��<0.001)������������СѪ�ܼ��Ͷ����ٷֱ����Ը��ڶ�����(q=14.57, P<0.01)������+L-Arg������СѪ�ܼ��Ͷ����ٷֱ����Ե��ڵ�����(q=9.19, P<0.01)�����Ը��ڶ�����(q=5.38, P<0.01)������������СѪ�ܷǼ���Ѫ�ܰٷֱ����Ե��ڶ�����(q=17.99,P<0.01), ����+L-Arg������СѪ�ܷǼ���Ѫ�ܰٷֱ����Ը��ڵ�����(q=8.74,P<0.01)�����Ե��ڶ�����(q=9.19, P<0.01)��
3.���С�С���Ͷ��������Ĥ��ȼ����(��1)�����С�С���Ͷ��������Ĥ��ȼ������3�����֮����������������(P��<0.001)�������������еȼ��Ͷ���RMT��RMA���Ը��ڶ�����(q=7.63��7.69, P��<0.01)����С���Ͷ���RMT��RMAҲ���Ը��ڶ�����(q=12.39,12.16, P��<0.01)������+L-Arg�������еȼ��Ͷ���RMT��RMA���Ե��ڵ�����(q=5.65, 5.65,P��<0.01)�������������Ȳ�������������(q=1.98��2.04, P��>0.05)����С���Ͷ���RMT��RMA���Ե��ڵ�����(q=8.09, 7.9, P��<0.01)�����Ը��ڶ�����(q=4.3, 4.25, P��<0.05)��
, ��Ĵҽҩ
4.���С�С���Ͷ�����Ƥϸ�����ṹ�������������С�С���Ͷ�����Ƥϸ�������ƽ���ڳ�Ѫ���ڱڣ��ṹ������ƽ����ϸ������С�����������ͣ�������������˿ռ�ݣ������������ܰ�(ͼ1)�����������ζ�����Ƥϸ���������ͣ�����ͻ���ǻ�����ı�ʽ������Ƥϸ�����ͣ������ڿɼ��������ݣ����������ͣ����������ţ�ƽ����ϸ������ʴ�ͬ�̶���ϳɱ���ת���������м�˿�����ܰ��٣�����������������������ϸ��������(ͼ2)������+L-Arg�����ζ�����Ƥϸ�������ƽ���ڳ�Ѫ���ڱڣ������ͳ̶Ƚϵ�������ᣬ�������Լ��٣�����ƽ����ϸ�������������ͣ��������Դ��ڹ��ɱ���(ͼ3)��
ͼ1 ���������μ��Ͷ�������Ƥϸ����ƽ���ṹ������ƽ����ϸ�����������ͣ�������������˿�����ܰ���ռ�� ��10 000
, http://www.100md.com
ͼ2 ���������μ��Ͷ�������Ƥϸ�����ͣ������ڼ��������ݡ�ƽ����ϸ���ʺϳɱ��ͣ������ڿɼ�������������壬������������� ��20 000
ͼ3 ����+L-Arg�����μ��Ͷ�������Ƥϸ����ƽ���������Լ��١�ƽ����ϸ������������ ��20 000
5.Ѫ��NO�ļ�Ӻ���(��1)��Ѫ��NO��Ӻ�����3�����֮���������������(P<0.001)�����������Ѫ��NO��Ӻ������Ե��ڶ�����(q=3.94, P<0.05)������+L-Arg�����Ѫ��NO��Ӻ������Ը��ڵ�����(q=5.22, P<0.05), ���������Ƚϲ�������������(q=1.33,P>0.05)��
6.�ζ�����Ƥϸ��ET-1 mRNA������������ַζ���(77%��7%)��Ƥϸ��ET-1 mRNA����������ź�(+)(ͼ4)����������(14%��7%)������+L-Arg��(4%��4%)����������ζ�����Ƥϸ����ʾET-1 mRNA���������ź�(+)��������зdz���������(q=9.32,10.86��P��<0.01)���������鼰����+L-Arg�����ζ�����Ƥϸ��ET-1 mRNA�����������������(q=1.54��P>0.05)��������(86%��7%)������+L-Arg��(96%��4%)����ַζ�����Ƥϸ����ET-1 mRNA����(ͼ5)�������������������ζ���(23%��7%)��Ƥϸ����ET-1 mRNA���������зdz���������(q=9.32, q=10.86��P��<0.01)���������鼰����+L-Arg�����ζ�����Ƥϸ��ET-1 mRNA�����������������(q =1.54��P>0.05)��
, ��Ĵҽҩ
ͼ4 ��������ζ�����1%��50%��Ƥϸ����ʾET-1 mRNA�ӽ������ź� DAB��ɫ����250
ͼ5 ����+L-Arg�����ζ�����Ƥϸ��ET-1 mRNA���������ܵ����� DAB��ɫ����400
����
�����Էζ�����ѹ���ٴ��ڶ��ķμ���������չ��������Ҫ�IJ�����������[1]�������Է�Ѫ�ܽṹ�ع������Ե����Էζ�����ѹ����Ҫ��������������Ҫ�����Ƿζ���ƽ����ϸ���������ʺ���Ĥ��������СѪ�ܼ������Լ�ϸ������ʵ�����[2]�����о����֣���ѹ����2�ܴ���ζ���ѹ���������ߣ���Ѫ�ܽṹ�ع����⾵�·�СѪ���м����̶�������ǿ�����С�С���Ͷ���RMT��RMA�������ӣ��羵�·ζ�����Ƥϸ���������ʺ����ͣ���Ĥƽ����ϸ���������ʴ���ͬ�̶ȵ���ϳɱ���ת����ͬʱ����2�ܺ����Ѫ��NO��Ӻ������Խ��ͣ���ET-1 mRNA����������ǿ����ʾ������NOˮƽ�Ľ��ͺ�ET-1����������ǿ���ܲ����˵����Է�Ѫ�ܽṹ�ع��������Էζ�����ѹ���γɡ��ɴ��������ܽ�NOˮƽ���ߺ�/��ET-1ˮƽ���Ϳ��Է��ε����Է�Ѫ�ܽṹ�ع���Ϊ�ٴ����Ƶ����Էζ�����ѹ�ṩ��һ��˼·��
, ��Ĵҽҩ
�о���������Դ�Ը���L-Arg��(��)NO���Ի�������Է�Ѫ�ܽṹ�ع��Լ������Էζ�����ѹ���γ�[3-5]���ٴ����ѿ�ʼӦ��NO����ķ��������Ƴ���̥��ѭ����̥�������ۺ����ȼ���������ȡ�ó�����Ч[11]����NO������øߣ������������ӣ�����һ���Ķ������ã�������������ٴ�Ӧ�á�
L-ArgΪNO�ϳɵ�ǰ�壬���������ں�NO��ø������NO���������Ӧ��L-Arg��Ԥ�����Է�Ѫ�ܽṹ�ع����о��õ�Խ��Խ������ӣ����������δ��ȫ���������ʵ��֤ʵ��NO��Ѫ��ƽ����ϸ������ֳ��ֱ����������[12]����ʾL-Arg��Ԥ�����Է�Ѫ�ܽṹ�ع��Ļ���֮һ������ͨ��NO��ֱ�����á���һ��������ʵ�黹��ʾ��NO�������꾲����Ƥϸ��ET-1�ĺϳ�����ڣ����ɽ���Ѫ��ET-1ˮƽ[6]�������������о����Ҳ��ʾ��NO �����Ե�������ζ���ET-1�����������������[7]����ET-1��Ŀǰ���ֵ�������ǿ����Ѫ�ܻ��Զ��ģ����Ҿ���ǿ�ҵĴ�Ѫ��ƽ������ֳ������[8]���ɴ˼����ʾL-Arg��Ԥ�����Է�Ѫ�ܽṹ�ع�����һ���ܻ�����ͨ���ٽ�NO�ϳɣ��Ӷ��Դٽ�Ѫ��ƽ����ϸ����ֳ����ET-1���������á�
, ��Ĵҽҩ
Ϊ��ȷ��һ���ڻ��ƵĿ����ԣ����ǽ�L-Arg�����Ե����Էζ�����ѹ�����Ѫ�����ͳ��ṹ��Ѫ��NO�ļ�Ӻ����Լ��ζ�����Ƥϸ��ET-1 mRNA�����Ӱ�������ͬ���о������������L-Argʹ�����Էζ�����ѹ����ζ���ѹ�����Խ��ͣ���СѪ�ܼ����̶����Լ��ᣬ���С�С���Ͷ���RMT��RMA���Խ��ͣ��ζ������ṹ�ĸı����Ի��⣻ͬʱL-Argʹ��������Ѫ��NO��Ӻ����������߲���ʹ��������ζ�����Ƥϸ��ET-1 mRMA�������������ơ���ʾL-Argͨ���ٽ�����NO���ɣ��Ӷ����Ʒζ�����Ƥϸ��ET-1 mRMA����Ե����Է�Ѫ�ܽṹ�ع��Լ������Էζ�����ѹ���γ�����Ҫ�ĵ������á�
������Ŀ��������Ȼ��ѧ����������Ŀ(39470735, 39870844)
�ο����ף�
1 �ž���, ����. ȱ���Էζ�����ѹ. ����������ҽ�ƴ�ѧ���й�Э��ҽ�ƴ�ѧ���ϳ�����, 1994. 79.
, http://www.100md.com
2 Vender RL. Chronic hypoxic pulmonary hypertension: cell biology to pathophysiology. Chest, 1994, 106: 236-243.
3 �뽨�⣬�ž���. ��Դ��һ���������ڴ�������Է�Ѫ�ܽṹ�ع�. �л�������־��1999��37��104-106.
4 �ֺ�, ��Ӣ��, ��Ϊ, ��. L-������������ƶ�ȱ������ζ���ѹ��Ѫ�ܽṹ�仯��Ӱ�� . �й�ҽѧ��ѧԺѧ�� , 1997 , 19 : 179-184.
5 Mitani Y, Maruyama K, Sakurai M. Prolonged administration of L-Arginine ameliorates chronic pulmonary hypertension and pulmonary vascular remodeling in rats. Circulation, 1997, 96: 689-697.
, ��Ĵҽҩ
6 Kourembanas S, Mcquillan LP, Leung GK. et al. Nitric oxide regulates the expression of vasoconstrictors and growth factors by vascular endothelium under both normoxia and hypoxia. J Clin Invest, 1993, 92: 99-104.
7 �ž���, ������, �Ա�, ��. һ������������ȱ������ζ�����Ƥ�ػ�������Ӱ�� . �л�ҽѧ��־ , 1997 , 77 : 263-265.
8 Janakidevi K, Fisher MA, Del Vecchio PJ, et al. Endothelin-1 stimulates DNA synthesis and proliferation of pulmonary artery smooth muscle cells. Am J Physiol, 1992, 263: C1295-C1301.
, http://www.100md.com
9 Barth PJ, Kimpel C, Roy S, et al. An improved mathematical approach for the assessment of the medial thickness of pulmonary arteries. Path Res Pract, 1993, 189: 567-576.
10 Hislop AA, Springall DR, Buttery LDK, et al. Abundance of endothelial nitric oxide synthase in newborn intrapulmonary arteries. Arch Dis Child, 1995, 73: F17-F21.
11 Roberts JD, Polaner DM, Lang P, et al. Inhaled nitric oxide reverses hypoxemia in persistent pulmonary hypertension of the newborns. Lancet, 1992, 340: 818-819.
12 Garg UC, Hassid A. Nitric oxide-generating vasodilators and 8-bromo-cyclic guanosine monophosphate inhibit mitogenesis and proferation of cultured rat vascular smooth muscle cells. J Clin Invest, 1989, 83: 1774-1777.
�ո����ڣ�1999-08-24, http://www.100md.com