诱导人T细胞对猪抗原免疫耐受的体外实验研究
作者:李彦 吴军 陈维佩 易绍萱 姜庆 陈希炜 张宁
单位:陈维佩 第三军医大学应用解剖与手术学教研室,重庆 400038;李彦 吴军 易绍萱 姜庆 陈希炜 张宁 第三军医大学烧伤研究所,重庆 400038
关键词:免疫耐受;T细胞发育;异种移植
中国免疫学杂志991004 摘 要 目的:探讨诱导人T细胞对猪抗原免疫耐受的状况。方法:通过建立人幼稚淋巴细胞与猪抗原递呈细胞混合培养模型,以淋巴细胞增殖实验及毒性实验、DNA分析、淋巴细胞表面标志检测等手段,观测了幼稚淋巴细胞分化、发育过程中对异种负调细胞疫苗初次接触与再次刺激的免疫应答状况的变化。结果:发现在体外,异种负调细胞疫苗能诱导人幼稚及成熟T细胞产生免疫耐受;辅助信号系统耐受在诱导过程中起着至关重要的作用。结论:提示在该系统中免疫耐受发生的机理是克隆排除与免疫无应答。
, 百拇医药 中国图书分类号 R392.11
An experimental study on the in vitro induced immune tolerance of human
T cell to porcine antigen-presenting cells
LI Yan,WU Jun,CHEN Wei-Pei et al.
Institute of Burn Injury and Department of Applied Surgical Anatomy and Operation Surgery,The 3rd Military Medical University,Chongqing 400038
Abstract Objective:To investigate the possibility and mechanism of the tolerance of human T cell to porcine antigen-presenting cells,the present study tried to prepare a kind of down-regulating cell vaccine(DRCV) and establish an in vitro novel model of immune tolerance,i.e.,mixed immature lymphocyte-porcine APC reaction.Methods:Based on this model,the lymphocyte proliferation,cytotoxicity,apoptosis,immunohistochemistry and FACS were used for analyze of immature T-cell differentiation,the reaction of immature T-cell to both primary and secondary APC stimulation,and the apoptosis of reactive T cells.Results:The results showed that:DRCV presented a very low level of CD86 expression,but the normal level of MHC-Ⅱ;DRCV could induce the antigen-specific immune tolerance of both immature and mature T cells and the underlying mechanism of this kind of immune tolerance was of anergy and clonal deletion occurred during the secondary encounter between human memorial T cells and the xeno-APC.Conclusion:According to the above data, propose that the expression of TCR on immature T-cell is regulated by antigens;the co-stimulatory signals play critical roles in the induction of both immature and matrue T-cell immune tolerance;and clonal deletion and unresponsiveness is responsible for the antigen-specific immune tolerance.
, 百拇医药
Key words Immune tolerance T cells Xeno-transplantation
近30年来,随着器官移植学的发展,特别是80年代以来,以CSA为代表的新型免疫抑制剂的出现以及多种单克隆抗体的广泛应用,有效地阻止和治疗了排斥综合症,使得同种异体移植的成功率显著提高[1],人类移植器官的来源缺乏成为主要矛盾[2]。由于猪的器官与人体器官大小形态相近,来源广泛,器官移植学上属于非协调性容易型移植,因此,选择以猪为供体来源具有极为现实的意义和广泛的应用前景[3]。但是,人对猪的排斥反应更难以控制,由于同时免疫抑制剂降低和破坏了机体正常的免疫防御体系,这些都是器官移植学急待解决的问题。免疫耐受是指针对内源性和外源性抗原特异性的长期免疫反应状态。所以,只有宿主产生对供体的免疫耐受,才是供体移植物长期存活于宿主体内而不被排斥的根本保证[4]。免疫耐受的诱导及其机理的研究是涉及基础理论及临床应用的重大课题。然而,目前所有的诱导方案均因各自的缺陷尚不能用于临床,免疫耐受的机理也远未阐明。本实验拟通过创建新的体外免疫耐受模型和制备异种负调细胞疫苗,采用淋巴细胞定向培养、淋巴细胞增殖实验、淋巴细胞毒性实验、DNA 分析、流式细胞仪检测细胞表面抗原和受体表达、免疫组化等手段,研究幼稚淋巴细胞在此模型中的分化与发育,发育过程中的淋巴细胞对异种负调细胞疫苗初次接触与再次刺激的免疫应答及其细胞凋亡等,探讨负调细胞疫苗诱导幼稚及成熟T细胞免疫耐受的可能性及相应的细胞和分子生物学机制,为寻找符合临床应用条件的有效诱导策略与方法提供理论和实践依据。
, 百拇医药
1 材料与方法
1.1 材料来源及分组 本实验分别采用健康人清除成熟T细胞的骨髓细胞作为耐受模型中受者免疫系统以及健康人外周血单个核细胞作为外周耐受实验中受者免疫系统。健康成年食用家猪外周血抗原递呈细胞-单核细胞作为异种供体刺激原(以下简称抗原细胞);将用1%多聚甲醛处理后的猪单核细胞作为耐受原-诱导疫苗(以下简称疫苗细胞-vaccine)[5]。Balb/c 小白鼠外周血的单核细胞作为第3种族来源的抗原。
模型分为3组:对照组(Control),疫苗诱导组(Group induced by vaccine,GIV),抗原刺激组(Group stimulated by porcine APC,GSA)。每组重复3个样本。各组经细胞培养15 d左右,检测相关指标。
相关试剂来源:IMDM培养基:美国Gibco公司。胎牛血清:美国Hylone公司。rhIL-3:第三军医大学防原教研室,10×104 U/支。CD2、CD3、CD4、CD8鼠抗人单克隆抗体:北京中山生物技术有限公司产品。新鲜AB型人血清:西南医院血库提供。免疫组化试剂(ABC):北京中山生物技术有限公司产品。D5637、FITC-CD2、FITC-CD3单克隆抗体:华西医科大学附属一院提供。P-I染液:华西医科大学附属一院提供,50 mg/ml。3H-TdR:中国同位素公司北京原子能研究所,1 mCi/m1。CytoTox96,Non-radioactive Assay 试剂盒:美国Promega公司。CD86(B7-2)单抗:美国Serotec公司。
, 百拇医药
1.2 实验模型的建立
1.2.1 完全培养液的制备 IMDM,10%胎牛血清,谷氨酰胺,2-巯基乙醇,硫酸庆大霉素,rhIL-3。
1.2.2 人幼稚骨髓单个核细胞的制备 取健康人骨髓单个核细胞,加入CD3单克隆抗体和人补体,清除成熟T细胞后备用。
1.2.3 人外周血单个核细胞的制备 取健康人外周血分离得到单个核细胞备用。
1.2.4 供体抗原及疫苗的制备 取新鲜猪外周血单核细胞作为异种供体细胞抗原。将一部分用1%多聚甲醛固定处理作为诱导疫苗。
1.2.5 骨髓细胞耐受诱导模型的建立 制备的供体抗原及疫苗按1:15比例分别加入1.2.2中备用的骨髓细胞中,组成抗原刺激组(GSA)和疫苗诱导组(GIV);加1 ml IMDM培养液于上述1.2.2中备用的骨髓细胞中组成对照组。于37℃ 5%CO2培养液中培养13~15 d,进行指标观察。
, 百拇医药
1.2.6 鼠抗原靶细胞的制备 Balb/c小鼠外周血单核细胞,2 000 rad 60 Co照射,调定浓度1×105 cells/ml,备用。
1.3 观测指标
1.3.1 观测骨髓细胞以及外周血细胞在本培养模型中的生长状况。
1.3.2 用流式细胞仪检测骨髓耐受诱导模型各组T细胞在生长过程中部分表面标志、CD2、CD3表面标志。
1.3.3 观测骨髓耐受诱导模型各组细胞生长过程中细胞周期与细胞凋亡。
1.3.4 用混合淋巴细胞增殖试验单向法(MLR)检测骨髓耐受诱导模型中各组接受再刺激的淋巴细胞增殖状况。
1.3.5 用细胞毒性试验检测骨髓耐受诱导模型中各组接受再刺激后的淋巴细胞的杀伤功能。
, 百拇医药
1.3.6 观测1%多聚甲醛固定后的单核细胞表面标志CD86(B7-2)及MHC Ⅱ表达。
1.4 数据处理 均以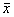 ±s表示,采用Microsoft Excel 97统计程序及本校SPMR程序包进行单因素、双因素方差分析。
±s表示,采用Microsoft Excel 97统计程序及本校SPMR程序包进行单因素、双因素方差分析。
2 结果
2.1 有关骨髓诱导模型的数据
2.1.1 各组骨髓细胞在此培养条件下的生长状况 见表1。在本培养体系中,CD3骨髓细胞在生长的第1天至第5天活细胞数明显下降,以后其数量逐步回升。第11~13天,细胞生长趋于稳定,两者比较,P>0.05,无显著差异。各实验组之间各个时相点的活细胞数基本一致,无显著差别(P>0.05)。
, 百拇医药
2.1.2 外周血单个核细胞在此完全培养条件下的生长状况 见表2。在本培养体系中,第1~5 天活细胞数明显下降,P<0.01,以后其数量进一步下降;5~9 d间比较,P<0.05;11 d以后大多死亡。
2.2 模型各组细胞生长过程中CD2、CD3的表达状况 见表3。各组骨髓细胞培养当天,清除成熟T细胞,流式细胞仪检测结果显示CD2+、CD3+细胞几乎无表达。经20 d培养,对照组、疫苗诱导组分别与培养当天各组比较,P<0.01,差异极显著,提示从幼稚淋巴细胞发育而来CD2+细胞有显著增加。对照组与疫苗组CD2表达的比较,P>0.05,无显著差异。
2.3 模型各组细胞生长过程中细胞周期与凋亡 见表4。在培养过程中,运用流式细胞仪观测骨髓耐受诱导模型中各组细胞Pre-G1期凋亡峰,测定细胞凋亡率。两两比较培养各组,P>0.05,它们之间无显著差异,各组值基本正常。
, 百拇医药
2.4 模型各组细胞接受再刺激的淋巴细胞增殖状况
2.4.1 各组接受同一猪抗原再刺激的淋巴细胞增殖状况 见表5。骨髓耐受诱导模型中各组培养15 d 后,分别与同一猪外周血单个核细胞的混合淋巴细胞反应单向法(MLR)的DPM值减去背景增殖而得出净增殖量,各组之间两两比较,P<0.01,存在极显著差异;抗原刺激组较其它两组增殖明显高,疫苗组淋巴细胞表现为抑制。
2.4.2 各组接受第3种族来源(鼠)抗原再刺激的淋巴细胞增殖状况 见表6。与以上相同培养条件下,各组细胞与鼠单个核细胞(2 000 rad irr)的MLR、DPM值基本一致。各组之间进行单因素方差分析,两两比较,P>0.05,它们之间无显著差异。
2.5 模型各组细胞接受同一抗原再刺激的细胞毒效应 见表7。培养15 d后,将各组细胞分别作为效应细胞,同一猪外周血单个核细胞作为靶细胞,以15:1的效靶比作用5 h,检测各组人淋巴细胞对同一猪抗原再接触时产生的细胞毒性杀伤效应变化。对照组、疫苗组的细胞毒性较小,而抗原组的毒性杀伤力很强。
, 百拇医药
2.6 诱导细胞与人外周血淋巴细胞混合培养的增殖状况 见表8。人外周血淋巴细胞对疫苗细胞(Vaccine cells,VC)初次接触和再次刺激反应以及猪抗原递呈细胞APC(单核细胞)初次反应之间两两比较,P>0.05,它们之间无显著差异。猪抗原细胞和人外周血淋巴细胞的再次反应与其它组相比较,P<0.05,存在显著差异,显示了抗原细胞再次反应较其它3组增殖效应明显高。
表1 各组不同时相点上的活细胞数(×103 cells/ml,±s,n=4)
Tab.1 The number of viable cells in the different groups at the growing phases(×103 cells/ml,±s,n=4) Group
, http://www.100md.com
1st day
5th day
7th day
10th day
13th day
Control1)
8502)
250±142)
315±712)
495±711)
530±281)
, 百拇医药
GIV1)
8502)
305±212)
365±712)
560±141)
470±141)
GSA1)
8502)
285±712)
345±712)
, 百拇医药
540±281)
505±351)
Note:1)P>0.05,2)P<0.01
表2 各组不同时相点上的活细胞数(×103 cells/ml,±s,n=4)
Tab.2 The number of viable cells in the different groups at the growing phases (×103 cells/ml,±s,n=4) 1st day
, 百拇医药
3rd day
5th day
7th day
9th day
11th day
500.0±0.01)
305.0±49.01)
58.0±2.81)
39.5±2.52)
34.0±9.62)
, http://www.100md.com 13.6±0.0
Note:1)P<0.01,2)P>0.05
表3 各组细胞CD2、CD3分子的阳性表达率
Tab.3 The expressing rate of CD2,CD3 in the groups Group
CD2+
CD3+
All the groups(at the beginning)
1.80%
0.70%
Control(has cultrued for 20 d)
, 百拇医药
14.60%
0.20%
GIV (has cultured for 20 d)
18.80%
12.40%
表4 各组细胞凋亡率(±s,n=3)
Tab.4 The apoptosis rate of the different groups(±s,n=3) Group
Apoptosis rate
, 百拇医药
Control
(6.13±0.23)%1)
GIV
(5.47±0.35)%1)
GSA
(5.97±0.31)%1)
Note:1)P>0.05
表5 同一抗原再刺激各组的MLR(±s,n=6)
Tab.5 MLR between the different groups and the same porcine antigen(x±s,n=6) Group
, http://www.100md.com
3H-TdR DPM
Control
39 091±8 5891)
GIV
-37 630±8 3211)
GSA
63 833±19 7031)
Note:1)P<0.01
表6 异种(鼠)抗原再刺激各组MLR(±s,n=5)
, 百拇医药
Tab.6 MLR between the different groups and mouse antigen(±s,n=5) Group
3H-TdR DPM
Control
130 209±61 8141)
GIV
102 044±17 3971)
GSA
119 227±14 9331)
, 百拇医药
Note:1)P>0.05
表7 各培养组细胞对同一抗原的杀伤率(n=3)
Tab.7 The cytotoxic ratio of the different groups to the same porcine cells(n=3) Group
Cytotoxic ratio
Control
13.09%
GIV
19.41%
GSA
100.00%
, http://www.100md.com
表8 人外周血对不同处理的猪抗原的MLR(单向法,±s,n=3)
Tab.8 MLR between the human peripheral lymphocyte and porcine APCs treated by different methods(±s,n=3) Group
3H-TdR DPM
Primary reaction with VC
6 919.3±1 053.31)
Primary reaction with APC
, 百拇医药
8 972.8±3 368.91)
Secondary reaction with VC
6 863.4±1 761.21)
Secondary reaction with APC
43 532.4±5 186.42)
Note:1)P>0.05,2)P<0.05 comparison with the other 3 groups separatly
表9 疫苗细胞和抗原细胞CD86和MHC Ⅱ表达的比较(±s,n=3)
, 百拇医药
Tab.9 Expressing comparison of CD86 and MHC II between the normal APC and the APC treated by 1% paraformaldehyde(±s,n=3)
Vaccine cells
Porcine APC
CD86(B7-2)
(3.67±1.05)%1)
(40.35±1.24)%1)
HLA-DR
, http://www.100md.com
(35.15±1.02)%2)
(37.81±1.33)%2)
Note:1)P<0.01,2)P>0.05
2.7 疫苗及猪抗原细胞部分表面标志的检测结果 见表9。检测显示两种细胞均有MHC II类抗原存在,它们之间的阳性表达率没有显著差别(P>0.5)。而疫苗细胞(VC)B7-2分子的表达较抗原细胞有显著缺乏(P<0.01)。
3 讨论
3.1 关于骨髓诱导模型对免疫耐受研究的作用与评价 本实验率先设计骨髓耐受诱导的体外模型。在淋巴系定向培养体系中[6,7],去除成熟T细胞的骨髓细胞培养5 d后,活细胞数量会有一个明显下降的过程,而后几天,细胞数量回升,说明有新生细胞产生。同时,外周中单个核细胞在此体系中培养,前5 d活细胞数量有相似的下降,而后几天,活细胞量会进一步减少,由此本培养条件适合骨髓细胞的生长而不适合外周血细胞的增殖生长,进而说明前5 d细胞的下降主要是较为成熟、无分裂增殖能力的细胞死亡或凋亡,而后的骨髓细胞中增多的群体来自有增殖分化能力的较为幼稚阶段的细胞群体。同时,开始有新的成熟细胞在几天后因缺乏维持其生长的条件而死亡,这可能在10 d以后,生长与死亡速度达到一个平衡,细胞数量相对保持稳定。此时通过流式细胞仪和免疫组化已证实新的来自于淋巴系祖细胞的CD2+细胞的产生以及一些CD3+细胞的产生,这些细胞的功能如何正是我们所要关注的。
, 百拇医药
3.2 受诱导而成熟的T细胞的DNA分析 将不同处理的猪抗原提呈细胞加入CD3-细胞的人骨髓细胞的培养体系中,经培养15 d左右,至有新生细胞的分化生长。通过DNA分析,了解这些新生细胞的Pre-G1期的凋亡峰无显著差别,显示没有克隆排除发生,提示了在异种耐受原与受者免疫系统初次接触阶段并非发生克隆排除,而是通过其它途径产生了免疫特殊记忆。
3.3 受诱导而成熟的T细胞的免疫效应的变化
3.3.1 识别与增殖的变化 混合淋巴细胞增殖反应单向法的检测,发现对照组中淋巴细胞与猪单个核细胞初次识别并表现一定量的增殖;而抗原刺激组与同一猪淋巴细胞再接触时,发生了更为明显的识别增殖,说明了CD3-的人骨髓细胞中有一些细胞对猪抗原细胞有较早的识别和记忆;特别发现为疫苗诱导后的T细胞在与同一猪的淋巴细胞再接触时表现为增殖的抑制。相对于对照组的初识别来看,疫苗组淋巴细胞的这种再次接触同样抗原(包括主要与次要组织相容性抗原)所表现的抑制是建立在识别基础上的免疫负调节作用而引起的,即首先是初识别、记忆,再识别,然后增殖受到抑制,因此可推测这种抑制效应是特异性的。
, http://www.100md.com
为了进一步证明这种效应的特异性,也可反证这种效应是建立在识别的基础上,通过鼠的外周血单核细胞作为第3来源的抗原细胞与各组培养的人幼稚淋巴细胞进行反应,发现各组均对该鼠抗原表现出相同的淋巴细胞的增殖反应。这就充分说明了在识别阶段上,疫苗诱导的骨髓细胞中成熟的T细胞对同一猪抗原的再接触具有再识别与负向调节作用,提示了再次接触相关抗原时发生了克隆排除,从而导致了免疫耐受发生。因此,可以确定免疫耐受发生不是单一步骤完成的。
3.3.2 各组免疫应答过程中效应阶段的特征与变化 以各培养组淋巴细胞为效应细胞,以同一猪的单核细胞作为靶细胞。结果表明对照组初次接触靶细胞仅产生13.0%的杀伤率,疫苗诱导组再次接触相同MHC的靶细胞后杀伤率只有19.4%,而抗原刺激组的细胞受到再次刺激时,表现出极强的细胞毒性效应,杀伤率达100.0%,证实了疫苗诱导下成熟的淋巴细胞(主要是T细胞)对同一猪抗原产生了免疫耐受。
3.4 本实验中细胞疫苗对外周血的免疫应答影响 本实验表明了人外周血细胞对疫苗细胞的MLR初次反应和再次反应没有明显的差别(P>0.05),其DPM值与猪抗原细胞的初次反应相似,而对猪抗原再刺激,则发生更为强烈的增殖(P<0.01),从而提示此疫苗对外周免疫细胞亦有耐受诱导的特征。
, 百拇医药
3.5 本实验中的细胞疫苗的分子结构的观察 通过对两种细胞表面MHC II抗原及B7-2分子表达状况的检测[8],结果证明疫苗细胞较完整地保留了供体MHC II抗原特性,而疫苗细胞表面B7-2分子的表达较供体单核细胞大为减少(P<0.01)。从而揭示本疫苗诱导免疫耐受的发生可能与共刺激信号B7-2分子表达缺乏有关。
资金项目:本课题受全军杰出人才基金资助
作者简介:李彦,男,28岁,博士生,讲师,从事器官移植免疫研究;
吴军,男,36岁,教授,博士生导师,从事器官移植免疫研究;
陈维佩,男,63岁,教授,硕士生导师,从事脾脏外科与脾移植研究
参考文献
, http://www.100md.com
1 Kabelitz D,da Silva Lobo M L,Pechhold K.Perspectives in transplantation immunology,1991.Klin Wochenschr,1991;69:623
2 Platt J L.Xenotransplantation:recent progress and current perspectives.Curr Opin Immunol,1996;8:721
3 Sachs D H.The pig as a potential xenograft donor.Vet Immunol Immunopathol,1994;43:185
4 Greenstein J L,Sachs D H.The use of tolerance for transplantation across xenogeneic barriers.Nat Biotechnol,1997;15:235
, 百拇医药
5 Sloan-Lancaster J,Evavold B D,Allen P M.Th2 cell clonal anergy as a consequence of partial activation.J Exp Med,1994;180:4,1195
6 Aiba Y,Mazda O,Davis M M et al.Requirement of a second signal from antigen presenting cells in the clonal deletion of immature T cells.Int Immunol,1994;6:1475
7 Chervenak R,Dempsey D,Soloiff R S et al.In vitro growth of bone marrow-resident T cell precursors supported by mast cell growth factor and IL-3.J Immunol,1992;149:2851
8 Anderson D E,Ausuberl L J,Krieger J et al.Weak peptide agonists reveal functional differences in B7-1 and B7-2 costimulation of human T cell clones.J Immunol,1997;159:1669
收稿1998-11-28 修回1999-01-29
, 百拇医药
单位:陈维佩 第三军医大学应用解剖与手术学教研室,重庆 400038;李彦 吴军 易绍萱 姜庆 陈希炜 张宁 第三军医大学烧伤研究所,重庆 400038
关键词:免疫耐受;T细胞发育;异种移植
中国免疫学杂志991004 摘 要 目的:探讨诱导人T细胞对猪抗原免疫耐受的状况。方法:通过建立人幼稚淋巴细胞与猪抗原递呈细胞混合培养模型,以淋巴细胞增殖实验及毒性实验、DNA分析、淋巴细胞表面标志检测等手段,观测了幼稚淋巴细胞分化、发育过程中对异种负调细胞疫苗初次接触与再次刺激的免疫应答状况的变化。结果:发现在体外,异种负调细胞疫苗能诱导人幼稚及成熟T细胞产生免疫耐受;辅助信号系统耐受在诱导过程中起着至关重要的作用。结论:提示在该系统中免疫耐受发生的机理是克隆排除与免疫无应答。
, 百拇医药 中国图书分类号 R392.11
An experimental study on the in vitro induced immune tolerance of human
T cell to porcine antigen-presenting cells
LI Yan,WU Jun,CHEN Wei-Pei et al.
Institute of Burn Injury and Department of Applied Surgical Anatomy and Operation Surgery,The 3rd Military Medical University,Chongqing 400038
Abstract Objective:To investigate the possibility and mechanism of the tolerance of human T cell to porcine antigen-presenting cells,the present study tried to prepare a kind of down-regulating cell vaccine(DRCV) and establish an in vitro novel model of immune tolerance,i.e.,mixed immature lymphocyte-porcine APC reaction.Methods:Based on this model,the lymphocyte proliferation,cytotoxicity,apoptosis,immunohistochemistry and FACS were used for analyze of immature T-cell differentiation,the reaction of immature T-cell to both primary and secondary APC stimulation,and the apoptosis of reactive T cells.Results:The results showed that:DRCV presented a very low level of CD86 expression,but the normal level of MHC-Ⅱ;DRCV could induce the antigen-specific immune tolerance of both immature and mature T cells and the underlying mechanism of this kind of immune tolerance was of anergy and clonal deletion occurred during the secondary encounter between human memorial T cells and the xeno-APC.Conclusion:According to the above data, propose that the expression of TCR on immature T-cell is regulated by antigens;the co-stimulatory signals play critical roles in the induction of both immature and matrue T-cell immune tolerance;and clonal deletion and unresponsiveness is responsible for the antigen-specific immune tolerance.
, 百拇医药
Key words Immune tolerance T cells Xeno-transplantation
近30年来,随着器官移植学的发展,特别是80年代以来,以CSA为代表的新型免疫抑制剂的出现以及多种单克隆抗体的广泛应用,有效地阻止和治疗了排斥综合症,使得同种异体移植的成功率显著提高[1],人类移植器官的来源缺乏成为主要矛盾[2]。由于猪的器官与人体器官大小形态相近,来源广泛,器官移植学上属于非协调性容易型移植,因此,选择以猪为供体来源具有极为现实的意义和广泛的应用前景[3]。但是,人对猪的排斥反应更难以控制,由于同时免疫抑制剂降低和破坏了机体正常的免疫防御体系,这些都是器官移植学急待解决的问题。免疫耐受是指针对内源性和外源性抗原特异性的长期免疫反应状态。所以,只有宿主产生对供体的免疫耐受,才是供体移植物长期存活于宿主体内而不被排斥的根本保证[4]。免疫耐受的诱导及其机理的研究是涉及基础理论及临床应用的重大课题。然而,目前所有的诱导方案均因各自的缺陷尚不能用于临床,免疫耐受的机理也远未阐明。本实验拟通过创建新的体外免疫耐受模型和制备异种负调细胞疫苗,采用淋巴细胞定向培养、淋巴细胞增殖实验、淋巴细胞毒性实验、DNA 分析、流式细胞仪检测细胞表面抗原和受体表达、免疫组化等手段,研究幼稚淋巴细胞在此模型中的分化与发育,发育过程中的淋巴细胞对异种负调细胞疫苗初次接触与再次刺激的免疫应答及其细胞凋亡等,探讨负调细胞疫苗诱导幼稚及成熟T细胞免疫耐受的可能性及相应的细胞和分子生物学机制,为寻找符合临床应用条件的有效诱导策略与方法提供理论和实践依据。
, 百拇医药
1 材料与方法
1.1 材料来源及分组 本实验分别采用健康人清除成熟T细胞的骨髓细胞作为耐受模型中受者免疫系统以及健康人外周血单个核细胞作为外周耐受实验中受者免疫系统。健康成年食用家猪外周血抗原递呈细胞-单核细胞作为异种供体刺激原(以下简称抗原细胞);将用1%多聚甲醛处理后的猪单核细胞作为耐受原-诱导疫苗(以下简称疫苗细胞-vaccine)[5]。Balb/c 小白鼠外周血的单核细胞作为第3种族来源的抗原。
模型分为3组:对照组(Control),疫苗诱导组(Group induced by vaccine,GIV),抗原刺激组(Group stimulated by porcine APC,GSA)。每组重复3个样本。各组经细胞培养15 d左右,检测相关指标。
相关试剂来源:IMDM培养基:美国Gibco公司。胎牛血清:美国Hylone公司。rhIL-3:第三军医大学防原教研室,10×104 U/支。CD2、CD3、CD4、CD8鼠抗人单克隆抗体:北京中山生物技术有限公司产品。新鲜AB型人血清:西南医院血库提供。免疫组化试剂(ABC):北京中山生物技术有限公司产品。D5637、FITC-CD2、FITC-CD3单克隆抗体:华西医科大学附属一院提供。P-I染液:华西医科大学附属一院提供,50 mg/ml。3H-TdR:中国同位素公司北京原子能研究所,1 mCi/m1。CytoTox96,Non-radioactive Assay 试剂盒:美国Promega公司。CD86(B7-2)单抗:美国Serotec公司。
, 百拇医药
1.2 实验模型的建立
1.2.1 完全培养液的制备 IMDM,10%胎牛血清,谷氨酰胺,2-巯基乙醇,硫酸庆大霉素,rhIL-3。
1.2.2 人幼稚骨髓单个核细胞的制备 取健康人骨髓单个核细胞,加入CD3单克隆抗体和人补体,清除成熟T细胞后备用。
1.2.3 人外周血单个核细胞的制备 取健康人外周血分离得到单个核细胞备用。
1.2.4 供体抗原及疫苗的制备 取新鲜猪外周血单核细胞作为异种供体细胞抗原。将一部分用1%多聚甲醛固定处理作为诱导疫苗。
1.2.5 骨髓细胞耐受诱导模型的建立 制备的供体抗原及疫苗按1:15比例分别加入1.2.2中备用的骨髓细胞中,组成抗原刺激组(GSA)和疫苗诱导组(GIV);加1 ml IMDM培养液于上述1.2.2中备用的骨髓细胞中组成对照组。于37℃ 5%CO2培养液中培养13~15 d,进行指标观察。
, 百拇医药
1.2.6 鼠抗原靶细胞的制备 Balb/c小鼠外周血单核细胞,2 000 rad 60 Co照射,调定浓度1×105 cells/ml,备用。
1.3 观测指标
1.3.1 观测骨髓细胞以及外周血细胞在本培养模型中的生长状况。
1.3.2 用流式细胞仪检测骨髓耐受诱导模型各组T细胞在生长过程中部分表面标志、CD2、CD3表面标志。
1.3.3 观测骨髓耐受诱导模型各组细胞生长过程中细胞周期与细胞凋亡。
1.3.4 用混合淋巴细胞增殖试验单向法(MLR)检测骨髓耐受诱导模型中各组接受再刺激的淋巴细胞增殖状况。
1.3.5 用细胞毒性试验检测骨髓耐受诱导模型中各组接受再刺激后的淋巴细胞的杀伤功能。
, 百拇医药
1.3.6 观测1%多聚甲醛固定后的单核细胞表面标志CD86(B7-2)及MHC Ⅱ表达。
1.4 数据处理 均以
±s表示,采用Microsoft Excel 97统计程序及本校SPMR程序包进行单因素、双因素方差分析。2 结果
2.1 有关骨髓诱导模型的数据
2.1.1 各组骨髓细胞在此培养条件下的生长状况 见表1。在本培养体系中,CD3骨髓细胞在生长的第1天至第5天活细胞数明显下降,以后其数量逐步回升。第11~13天,细胞生长趋于稳定,两者比较,P>0.05,无显著差异。各实验组之间各个时相点的活细胞数基本一致,无显著差别(P>0.05)。
, 百拇医药
2.1.2 外周血单个核细胞在此完全培养条件下的生长状况 见表2。在本培养体系中,第1~5 天活细胞数明显下降,P<0.01,以后其数量进一步下降;5~9 d间比较,P<0.05;11 d以后大多死亡。
2.2 模型各组细胞生长过程中CD2、CD3的表达状况 见表3。各组骨髓细胞培养当天,清除成熟T细胞,流式细胞仪检测结果显示CD2+、CD3+细胞几乎无表达。经20 d培养,对照组、疫苗诱导组分别与培养当天各组比较,P<0.01,差异极显著,提示从幼稚淋巴细胞发育而来CD2+细胞有显著增加。对照组与疫苗组CD2表达的比较,P>0.05,无显著差异。
2.3 模型各组细胞生长过程中细胞周期与凋亡 见表4。在培养过程中,运用流式细胞仪观测骨髓耐受诱导模型中各组细胞Pre-G1期凋亡峰,测定细胞凋亡率。两两比较培养各组,P>0.05,它们之间无显著差异,各组值基本正常。
, 百拇医药
2.4 模型各组细胞接受再刺激的淋巴细胞增殖状况
2.4.1 各组接受同一猪抗原再刺激的淋巴细胞增殖状况 见表5。骨髓耐受诱导模型中各组培养15 d 后,分别与同一猪外周血单个核细胞的混合淋巴细胞反应单向法(MLR)的DPM值减去背景增殖而得出净增殖量,各组之间两两比较,P<0.01,存在极显著差异;抗原刺激组较其它两组增殖明显高,疫苗组淋巴细胞表现为抑制。
2.4.2 各组接受第3种族来源(鼠)抗原再刺激的淋巴细胞增殖状况 见表6。与以上相同培养条件下,各组细胞与鼠单个核细胞(2 000 rad irr)的MLR、DPM值基本一致。各组之间进行单因素方差分析,两两比较,P>0.05,它们之间无显著差异。
2.5 模型各组细胞接受同一抗原再刺激的细胞毒效应 见表7。培养15 d后,将各组细胞分别作为效应细胞,同一猪外周血单个核细胞作为靶细胞,以15:1的效靶比作用5 h,检测各组人淋巴细胞对同一猪抗原再接触时产生的细胞毒性杀伤效应变化。对照组、疫苗组的细胞毒性较小,而抗原组的毒性杀伤力很强。
, 百拇医药
2.6 诱导细胞与人外周血淋巴细胞混合培养的增殖状况 见表8。人外周血淋巴细胞对疫苗细胞(Vaccine cells,VC)初次接触和再次刺激反应以及猪抗原递呈细胞APC(单核细胞)初次反应之间两两比较,P>0.05,它们之间无显著差异。猪抗原细胞和人外周血淋巴细胞的再次反应与其它组相比较,P<0.05,存在显著差异,显示了抗原细胞再次反应较其它3组增殖效应明显高。
表1 各组不同时相点上的活细胞数(×103 cells/ml,
±s,n=4)Tab.1 The number of viable cells in the different groups at the growing phases(×103 cells/ml,
±s,n=4) Group, http://www.100md.com
1st day
5th day
7th day
10th day
13th day
Control1)
8502)
250±142)
315±712)
495±711)
530±281)
, 百拇医药
GIV1)
8502)
305±212)
365±712)
560±141)
470±141)
GSA1)
8502)
285±712)
345±712)
, 百拇医药
540±281)
505±351)
Note:1)P>0.05,2)P<0.01
表2 各组不同时相点上的活细胞数(×103 cells/ml,
±s,n=4)Tab.2 The number of viable cells in the different groups at the growing phases (×103 cells/ml,
±s,n=4) 1st day, 百拇医药
3rd day
5th day
7th day
9th day
11th day
500.0±0.01)
305.0±49.01)
58.0±2.81)
39.5±2.52)
34.0±9.62)
, http://www.100md.com 13.6±0.0
Note:1)P<0.01,2)P>0.05
表3 各组细胞CD2、CD3分子的阳性表达率
Tab.3 The expressing rate of CD2,CD3 in the groups Group
CD2+
CD3+
All the groups(at the beginning)
1.80%
0.70%
Control(has cultrued for 20 d)
, 百拇医药
14.60%
0.20%
GIV (has cultured for 20 d)
18.80%
12.40%
表4 各组细胞凋亡率(
±s,n=3)Tab.4 The apoptosis rate of the different groups(
±s,n=3) GroupApoptosis rate
, 百拇医药
Control
(6.13±0.23)%1)
GIV
(5.47±0.35)%1)
GSA
(5.97±0.31)%1)
Note:1)P>0.05
表5 同一抗原再刺激各组的MLR(
±s,n=6)Tab.5 MLR between the different groups and the same porcine antigen(x±s,n=6) Group
, http://www.100md.com
3H-TdR DPM
Control
39 091±8 5891)
GIV
-37 630±8 3211)
GSA
63 833±19 7031)
Note:1)P<0.01
表6 异种(鼠)抗原再刺激各组MLR(
±s,n=5), 百拇医药
Tab.6 MLR between the different groups and mouse antigen(
±s,n=5) Group3H-TdR DPM
Control
130 209±61 8141)
GIV
102 044±17 3971)
GSA
119 227±14 9331)
, 百拇医药
Note:1)P>0.05
表7 各培养组细胞对同一抗原的杀伤率(n=3)
Tab.7 The cytotoxic ratio of the different groups to the same porcine cells(n=3) Group
Cytotoxic ratio
Control
13.09%
GIV
19.41%
GSA
100.00%
, http://www.100md.com
表8 人外周血对不同处理的猪抗原的MLR(单向法,
±s,n=3)Tab.8 MLR between the human peripheral lymphocyte and porcine APCs treated by different methods(
±s,n=3) Group3H-TdR DPM
Primary reaction with VC
6 919.3±1 053.31)
Primary reaction with APC
, 百拇医药
8 972.8±3 368.91)
Secondary reaction with VC
6 863.4±1 761.21)
Secondary reaction with APC
43 532.4±5 186.42)
Note:1)P>0.05,2)P<0.05 comparison with the other 3 groups separatly
表9 疫苗细胞和抗原细胞CD86和MHC Ⅱ表达的比较(
±s,n=3), 百拇医药
Tab.9 Expressing comparison of CD86 and MHC II between the normal APC and the APC treated by 1% paraformaldehyde(
±s,n=3)Vaccine cells
Porcine APC
CD86(B7-2)
(3.67±1.05)%1)
(40.35±1.24)%1)
HLA-DR
, http://www.100md.com
(35.15±1.02)%2)
(37.81±1.33)%2)
Note:1)P<0.01,2)P>0.05
2.7 疫苗及猪抗原细胞部分表面标志的检测结果 见表9。检测显示两种细胞均有MHC II类抗原存在,它们之间的阳性表达率没有显著差别(P>0.5)。而疫苗细胞(VC)B7-2分子的表达较抗原细胞有显著缺乏(P<0.01)。
3 讨论
3.1 关于骨髓诱导模型对免疫耐受研究的作用与评价 本实验率先设计骨髓耐受诱导的体外模型。在淋巴系定向培养体系中[6,7],去除成熟T细胞的骨髓细胞培养5 d后,活细胞数量会有一个明显下降的过程,而后几天,细胞数量回升,说明有新生细胞产生。同时,外周中单个核细胞在此体系中培养,前5 d活细胞数量有相似的下降,而后几天,活细胞量会进一步减少,由此本培养条件适合骨髓细胞的生长而不适合外周血细胞的增殖生长,进而说明前5 d细胞的下降主要是较为成熟、无分裂增殖能力的细胞死亡或凋亡,而后的骨髓细胞中增多的群体来自有增殖分化能力的较为幼稚阶段的细胞群体。同时,开始有新的成熟细胞在几天后因缺乏维持其生长的条件而死亡,这可能在10 d以后,生长与死亡速度达到一个平衡,细胞数量相对保持稳定。此时通过流式细胞仪和免疫组化已证实新的来自于淋巴系祖细胞的CD2+细胞的产生以及一些CD3+细胞的产生,这些细胞的功能如何正是我们所要关注的。
, 百拇医药
3.2 受诱导而成熟的T细胞的DNA分析 将不同处理的猪抗原提呈细胞加入CD3-细胞的人骨髓细胞的培养体系中,经培养15 d左右,至有新生细胞的分化生长。通过DNA分析,了解这些新生细胞的Pre-G1期的凋亡峰无显著差别,显示没有克隆排除发生,提示了在异种耐受原与受者免疫系统初次接触阶段并非发生克隆排除,而是通过其它途径产生了免疫特殊记忆。
3.3 受诱导而成熟的T细胞的免疫效应的变化
3.3.1 识别与增殖的变化 混合淋巴细胞增殖反应单向法的检测,发现对照组中淋巴细胞与猪单个核细胞初次识别并表现一定量的增殖;而抗原刺激组与同一猪淋巴细胞再接触时,发生了更为明显的识别增殖,说明了CD3-的人骨髓细胞中有一些细胞对猪抗原细胞有较早的识别和记忆;特别发现为疫苗诱导后的T细胞在与同一猪的淋巴细胞再接触时表现为增殖的抑制。相对于对照组的初识别来看,疫苗组淋巴细胞的这种再次接触同样抗原(包括主要与次要组织相容性抗原)所表现的抑制是建立在识别基础上的免疫负调节作用而引起的,即首先是初识别、记忆,再识别,然后增殖受到抑制,因此可推测这种抑制效应是特异性的。
, http://www.100md.com
为了进一步证明这种效应的特异性,也可反证这种效应是建立在识别的基础上,通过鼠的外周血单核细胞作为第3来源的抗原细胞与各组培养的人幼稚淋巴细胞进行反应,发现各组均对该鼠抗原表现出相同的淋巴细胞的增殖反应。这就充分说明了在识别阶段上,疫苗诱导的骨髓细胞中成熟的T细胞对同一猪抗原的再接触具有再识别与负向调节作用,提示了再次接触相关抗原时发生了克隆排除,从而导致了免疫耐受发生。因此,可以确定免疫耐受发生不是单一步骤完成的。
3.3.2 各组免疫应答过程中效应阶段的特征与变化 以各培养组淋巴细胞为效应细胞,以同一猪的单核细胞作为靶细胞。结果表明对照组初次接触靶细胞仅产生13.0%的杀伤率,疫苗诱导组再次接触相同MHC的靶细胞后杀伤率只有19.4%,而抗原刺激组的细胞受到再次刺激时,表现出极强的细胞毒性效应,杀伤率达100.0%,证实了疫苗诱导下成熟的淋巴细胞(主要是T细胞)对同一猪抗原产生了免疫耐受。
3.4 本实验中细胞疫苗对外周血的免疫应答影响 本实验表明了人外周血细胞对疫苗细胞的MLR初次反应和再次反应没有明显的差别(P>0.05),其DPM值与猪抗原细胞的初次反应相似,而对猪抗原再刺激,则发生更为强烈的增殖(P<0.01),从而提示此疫苗对外周免疫细胞亦有耐受诱导的特征。
, 百拇医药
3.5 本实验中的细胞疫苗的分子结构的观察 通过对两种细胞表面MHC II抗原及B7-2分子表达状况的检测[8],结果证明疫苗细胞较完整地保留了供体MHC II抗原特性,而疫苗细胞表面B7-2分子的表达较供体单核细胞大为减少(P<0.01)。从而揭示本疫苗诱导免疫耐受的发生可能与共刺激信号B7-2分子表达缺乏有关。
资金项目:本课题受全军杰出人才基金资助
作者简介:李彦,男,28岁,博士生,讲师,从事器官移植免疫研究;
吴军,男,36岁,教授,博士生导师,从事器官移植免疫研究;
陈维佩,男,63岁,教授,硕士生导师,从事脾脏外科与脾移植研究
参考文献
, http://www.100md.com
1 Kabelitz D,da Silva Lobo M L,Pechhold K.Perspectives in transplantation immunology,1991.Klin Wochenschr,1991;69:623
2 Platt J L.Xenotransplantation:recent progress and current perspectives.Curr Opin Immunol,1996;8:721
3 Sachs D H.The pig as a potential xenograft donor.Vet Immunol Immunopathol,1994;43:185
4 Greenstein J L,Sachs D H.The use of tolerance for transplantation across xenogeneic barriers.Nat Biotechnol,1997;15:235
, 百拇医药
5 Sloan-Lancaster J,Evavold B D,Allen P M.Th2 cell clonal anergy as a consequence of partial activation.J Exp Med,1994;180:4,1195
6 Aiba Y,Mazda O,Davis M M et al.Requirement of a second signal from antigen presenting cells in the clonal deletion of immature T cells.Int Immunol,1994;6:1475
7 Chervenak R,Dempsey D,Soloiff R S et al.In vitro growth of bone marrow-resident T cell precursors supported by mast cell growth factor and IL-3.J Immunol,1992;149:2851
8 Anderson D E,Ausuberl L J,Krieger J et al.Weak peptide agonists reveal functional differences in B7-1 and B7-2 costimulation of human T cell clones.J Immunol,1997;159:1669
收稿1998-11-28 修回1999-01-29
, 百拇医药