大鼠脑缺血再灌注后线粒体呼吸链相关基因表达
作者:袁有忠 吴琦 邓小明 熊源长 刘树孝
单位:袁有忠(长海医院血液科,上海 200433);吴琦 邓小明 熊源长 刘树孝(上海第二军医大学附属长海医院麻醉科, 上海 200433)
关键词:局部缺血;再灌注损伤;基因表达;DNA,线粒体;大鼠
中国病理生理杂志000813
[摘 要] 目的:研究海马神经元缺血再灌注后线粒体相关基因的表达。方法:改良Pulsinell四血管阻断模型致大鼠脑缺血10min再灌注24 h后,取新鲜海马组织抽提mRNA。逆转录后经液相杂交递减无差异表达序列。以抑制性PCR方式扩增差异表达序列。之后构建cDNA文库,测序鉴定阳性克隆。最后应用斑点杂交确认差异表达。结果:构建cDNA文库后获得的78个阳性克隆中有7个克隆经最后的鉴定具有差异表达性。测序提示为细胞色素氧化酶C亚单位和NADH-泛醌氧化酶复合体4链的表达性片段。结论:细胞色素氧化酶C亚单位的差异表达从另一个侧面证实了之前提出的再灌注损伤中细胞色素氧化酶C的重要作用。再灌注损伤后NADH-泛醌氧化酶复合体4链的表达提示原来功能不明的NADH-泛醌氧化酶复合体β亚基可能在这一病理过程中发挥一定的作用。
, 百拇医药
[中图分类号] R743.31 [文献标识码] A
[文章编号] 1000-4718(2000)08-0722-04
Mitochondria respiratory chain related genes expression
in post ischemia/reperfusion rat
WU Qi, DENG Xiao-ming, XIONG Yuan-chang, LIU Shu-xiao
(Department of Anesthesiology, The Second Military Medical University Affiliated Changhai Hospital, Shanghai 200433,China)
, 百拇医药
YUAN You-zhong
(Department of Hematology, The Second Military Medical University Affiliated Changhai Hospital, Shanghai 200433,China)
[Abstrac] AIM: Apply the newly invented method of suppression subtractive hybridization to scan the mitochondria genes expression of hippocampus neurons in post ischemia/reperfusion rats. METHODS: Decapitate the rats suffering 10 min of whole brain ischemia and 24 h of reperfusion in Pulsinelli’s 4 vessels occlusion ischemia/reperfusion model. Dissociated hippocampus and isolated mRNA. After reversed transcription, use SSH to subtract the common sequences, then use suppressed PCR to amplify the differentially expressed sequences. After cloning and DNA sequencing, the dot hybridization was used to finally identify the differential gene expression. RESULTS: In the 78 positive clones acquired from cloning, 7 clones represent differential gene expression after confirmed by clone PCR, dot blot and DNA sequencing. The DNA sequencing data indicate these clones are ESTs of Cytochrome c oxidase subunit and NADH-ubiquinone oxidoreductase chain 4 gene. CONCLUSION: Cytochrome C oxidase mRNA level changs during brain ischemia /reperfusion. The differential expression of NADH-ubiquinone oxidoreductase chain 4, one of those 13 subunits of subcomplex β, suggests that subcomplex β may have important function associated with pathophysiologic changs in reperfusion injury.
, 百拇医药
[MeSH] Ischemia;Reperfusion injuries;Gene expression;DNA,mitochondrial;Rats
脑缺血/再灌注损伤后常会观察到原有的一些神经功能损害程度并不是静止在缺血的当时,而是进行性加重。这是因为调节这些功能的神经元发生延迟性死亡了。这种病理变化通常是凋亡。发生凋亡的神经元在受到致死性刺激的最初48 h内还能保持结构的完整。这提示:凋亡是一个过程,如果在这时间之内能有一些有效措施阻止凋亡的级联效应,那么挽救凋亡神经元是可能的。凋亡还是一种耗能的程序性死亡。线粒体不仅因为它是能量转换器而涉及到凋亡的调控中,更有证据表明线粒体的某些组份直接参与了凋亡的调控过程[1]。Diatchenko等[2]报道的抑制递减杂交法(suppression subtractive hybridization, SSH)能有效检测两个不同细胞群差异表达基因的方法。本研究应用该方法检测缺血再灌注后线粒体基因的表达。
, 百拇医药
材 料 和 方 法
一、脑缺血模型复制与分组
选用雄性SD大鼠,体重250~300 g。脑缺血模型采用改良的Pulsinelli法复制,即异戊巴比妥麻醉下烧闭双侧椎动脉同时双侧颈总动脉套线。24 h后,动物清醒下,拉紧双侧缝线并保持一定张力,阻断双侧颈总动脉造成脑缺血10 min。剪断缝线并抽出,开放颈总动脉血流造成再灌注24 h。正常对照组与脑缺血组动物各6只,断头处死大鼠,打开颅腔,取出海马组织后迅速放入液氮中速冻。24 h后转移至-85℃低温冰箱中保存。
二、海马组织mRNA的抽提、鉴定和浓缩
液氮冷冻条件下将海马组织捣碎成粉末。应用组织/细胞总RNA抽提试剂盒(Watson公司)抽提总RNA。随后应用mRNA Isolation Kit for Blood/Bone Marrow(Boehringer-mannheim,Germany)抽提mRNA,即polyA(+)RNA。
, 百拇医药
三、抑制性递减杂交分析
未经特别说明,试剂主要由PCR-SelectTM cDNA Subtraction kit(Clontech, USA)提供。
cDNA合成引物:5′-TTTTGTACAAGCTT30N1N-3′。
连接器1序列: 5′-CTAATACGACTCACTATAGGGCTCGAGCGGCCGCCCGGGCAGGT-3′
3′-GGCCCGTCCA-5′
连接器2序列: 5′-CTAATACGACTCACTATAGGGCAGCGTGGTCGCGGCCGAGGT-3′
3′-GCCGGCTCCA-5′
, 百拇医药
PCR引物1: 5′-CTAATACGACTCACTATAGGGCTC-3′
巢式PCR引物1: 5′-TCGAGCGGCCGCCCGGGCAGGT-3′
巢式PCR引物2: 5′-AGCGTGGTCGCGGCCGAGGT -3′
(一)mRNA逆转录合成cDNA 模板采用经分光光度计精确定量的mRNA (0.5 μg/μL)。
(二)限制性酶切反应及连接器的连接反应 cDNA产物纯化后经RsaⅠ限制性酶切后,实验组酶切产物分成相同的两组,分别接上连接器1和连接器2。
(三)液相杂交 第一轮杂交是由两组独立的杂交反应组成的, 杂交体系的构成为RsaⅠ限制性酶切后的对照组cDNA,带连接器的实验组cDNA(两组杂交反应分别含不同的连接器),1×杂交缓冲液。第一轮杂交的反应参数(98℃变性1.5 min, 68 ℃保温8 h)。第一轮杂交后进入第二轮杂交反应。在同一杂交体系中混合上述两组独立的杂交反应。同时在杂交体系中加入新鲜变性的RsaⅠ限制性酶切后的对照组cDNA,造成杂交体系中该成分过量。第二轮杂交的反应参数(68℃杂交过夜)。
, 百拇医药
(四)抑制PCR 第一轮PCR体系中,模板为第二轮杂交产物,PCR引物为PCR引物1。第二轮PCR体系中,模板为第一轮PCR产物,PCR引物为巢式PCR引物1和巢式PCR引物2。
四、cDNA文库的构建和测序分析
利用T/A cloning kit(Promega,USA),将第二轮PCR扩增产物插入T-Vector中。根据α 互补的原理,在涂布X-gal的琼脂平板上筛选出带插入片断的菌落。进一步以菌落PCR鉴定。抽提质粒进行全自动DNA测序。质粒的抽提采用Plasmid Mini Kit(Qiagen,USA)。 通过e-mail与genbank联系,借助BLAST程序分析核酸及蛋白的同源性。
五、斑点杂交
将送测序的质粒中的插入片断制备成[α-32P]dATP标记的探针,[α-32P]dATP购自北京亚辉生物医学工程公司,探针标记的试剂来自MegaprimeTM DNA labelling systems(Life science,USA)。将大约1 μg正常组和实验组总RNA直接烤膜固定到尼龙膜上(Hybrond, Germany)。杂交过程和洗膜过程参照《分子克隆实验指南》[3]。杂交膜在cyclone磷屏成像系统中曝光、扫描成像后储存于计算机中。
, 百拇医药
结 果
初步构建了大鼠脑缺血再灌注后差异表达的cDNA 文库。根据α互补的原理筛选出78个阳性克隆。同时由于差异表达cDNA 在构建文库前已被RsaⅠ限制性酶切,片断大小约为0.1~2 kb。 因而在菌落PCR中可借助来进一步鉴别阳性克隆。进一步筛选后得到12个阳性克隆(图1)。阳性克隆抽提质粒后行全自动测序分析。其中1、2、5、10、11、12号克隆对应细胞色素氧化酶C亚单位;3号克隆为ATP合成酶e链;4号克隆为NADH-泛醌氧化酶复合体4链(ND4);6号克隆为40S核糖体蛋白;7号克隆为硫糖蛋白-2;8号克隆为tRNA; 9号克隆为ATP合成酶a链。选择4、5、7、9号克隆,按材料方法中所述步骤进一步以斑点杂交验证差异表达性,结果4、5号克隆显示了差异表达(图2)。
NADH-泛醌氧化酶复合体4链的表达片段资料为:
3'TCACAGGAATATACTCAATATATATGATTATCACAAC
, 百拇医药
CCAACGAGGAAAACTAACCAGCCACATAAACAACCT
CCAACCTTCCCACACACGAGAATTAACACTCATAGCT
CTACACATTATTCCCCTCATACTATTAACAATCAACCC
TAAACTCATCACAGGCCTAACAATA 5'
细胞色素氧化酶C亚单位的的表达片段资料为:
3'TTGACTCCTACATAATCCCAACCAATGACCTAAAACC
AGGTGAACTTCGTCTATTAGAAGTTGATAATCGGGTA
GTCTTACCAATAGAACTTCCAATTCGTATACTAATCT
, http://www.100md.com
CATCCGAAGACGTCCTGCACTCATGAGCCATCCCTTC
ACTAGGGTTAAAAACCGACGCAATCCCCGGCCGCCTA
AACCAAGCTACTAGTCACATCAAACCGACCAGGTCTA
TTCTATGGCCAATGCTCTGAAATTTGCGGCTCAAATC
ACAGCTTCATACCCATTGTAC 5'
Fig 1 Positive clones selected by clone PCR
M: DNA marker(ΦX174DNA HaeⅢ digested fragment) ; lane 1~8,11,12~14 have almost the same size fragment, so they may represent one clone ; lane 9,10,15,16 have different size fragment, therefor probably represent different clone
, 百拇医药
图1 菌落PCR筛选出的阳性克隆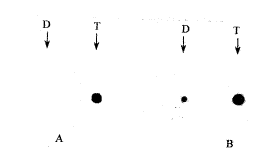
Fig 2 Dot hybridization of differential expressed clone
A: The 4th chain of NADH:ubiquinone oxidoreductase; B: Cytochrome C oxidase subunit; D: Control group; T: Test group
图2 差异表达克隆的斑点杂交分析
讨 论
试验采用的脑缺血模型是在Pulsinelli等创建的大鼠4条血管阻断所致全脑缺血模型的基础上进行的改进。我们曾观察该模型中阻断颈总动脉后局部脑血流量的变化,结果发现内侧中央前区、纹状皮层、小脑半球单小叶等脑区在脑缺血期间均显著下降,最低水平分别为缺血前的9%、11%、24%[4]。该模型造成的组织病理学变化、严重前脑缺血的重复性均良好,而且和经典Pulsinelli模型比较,阻断和开放颈总动脉系统更方便。
, http://www.100md.com
脑缺血再灌注后表现出差异表达的2个基因都是线粒体呼吸链的重要成分。 细胞色素氧化酶C(cytochrome C)位于线粒体内外膜间。它的功能是把电子从QH2-细胞色素氧化酶C还原酶复合体传递到细胞色素氧化酶C氧化酶复合体[5]。NADH-泛醌氧化酶复合体(NADH:ubiquinone oxidoreductase complex)也位于线粒体内外膜之间,它是电子进入呼吸链的第一道门户[6]。
线粒体长久以来是以能量转化中心的形象出现的,但最新的研究表明线粒体在细胞死亡的调控机制中发挥着重要的作用。在一项离体的细胞试验中发现,Bcl-2抑制核皱缩和DNA断裂的作用必须有线粒体参与[7]。但最早的突破是细胞色素氧化酶C注入培养细胞中引起凋亡表现的发现[8]。随后发现缺血再灌注损伤后细胞色素氧化酶C从线粒体中释放入胞浆中并和胱氨酸蛋白酶原-9(procaspase-9)和Apaf-1构成了“凋亡小体” ,进而激活其他半胱氨酸蛋白酶(caspase)共同完成一系列凋亡的生化过程[9,10]。还有学者观察到,大脑中动脉阻断90 min再灌注4 h, 细胞色素氧化酶C出现表达,再灌注24h达到高峰[11]。细胞色素氧化酶C的释放确切的机制不甚清楚。有研究提示线粒体内膜的通透性异常起了关键的作用。这种状态通常被定义为线粒体通透性转运(mitochondrial permeability transition, MPT),它允许分子量小于1500 D的分子自由扩散。当MPT成为不可逆变化后,线粒体内膜由于渗透压的作用极度皱缩,进而引起外膜破裂,细胞色素氧化酶C从内外膜间释放[12]。虽然线粒体外膜的破裂可以导致细胞色素氧化酶C、凋亡诱导因子(AIF)的释放,但是只有一部分线粒体的外膜破裂[13]。但有观察提示线粒体所有的细胞色素氧化酶C在凋亡时释放。这说明现有观察线粒体外膜破裂的手段不灵敏或者细胞色素氧化酶C的释放有其他机制[14]。
, 百拇医药
从牛心克隆到的NADH-泛醌氧化酶复合体由7个线粒体编码的亚基ND1- ND6(分子量50 kD左右)、ND4L和34个核编码的亚单位构成。对NADH-泛醌氧化酶复合体的结构功能分析显示:该复合体由 α、β亚单位构成;α亚单位含28个亚基,包括了全部的黄素蛋白和铁蛋白,集中了全部的具有还原活性的Fe-S簇,是NADH-泛醌氧化酶复合体氧化还原活性中心所在;β亚单位由13个亚基构成(包含ND4), 无氧化还原活性中心,其生化功能不明[15]。在NADH-泛醌氧化酶复合体中存在如此庞大、复杂的但却与主要功能无关的β亚单位,人们不禁怀疑它是否具有其他重要的生理功能?有学者提出凋亡调控过程中,MPT后线粒体外膜破裂,凋亡诱导因子(一类分子量50 kD左右的分子)从内外膜间释放,凋亡的下游过程启动[16]。NADH-泛醌氧化酶复合体4链(ND4)的分子量、定位、缺血再灌注后表达等特征,使之类似于凋亡诱导因子。但ND4和细胞死亡调控的确切关系需要进一步的严谨实验证实。
, 百拇医药
[参 考 文 献]
[1] Abe K, Aoki M, Kawagoe J, et al. Ischemic delayed neuronal death: A mitochondrial hypothesis[J]. Stroke, 1995, 26:1473~1489.
[2] Diatchenko L, Lau YFC, Campbell AP, et al. Suppression subtractive hybridization: A method for generating differentially regulated or tissue-specific cDNA probes and libraries[J]. Proc Natl Acad Sci, 1996, 93:6025~6030.
[3] J 萨姆布鲁克,EF弗里奇,T曼尼阿蒂斯著.分子克隆实验指南[M].第2版.北京:科学出版社,1996.65~68.
, http://www.100md.com
[4] 邓小明,石一鸣,王景阳,等.脑缺血再灌注损伤的动物模型[J].中国病理生理杂志, 1991, 7:208.
[5] Boyer PD, Chance B, Ernster L, et al. Oxidative phosphorylation and photo-phosphorylation[J]. Annu Rev Biochem, 1977, 46:955~1026.
[6] Friedrich T, Abelmann A, Brors B, et al. Redox components and structure of the respiratory NNDH:ubiquinone oxidoreductase(complexⅠ)[J]. Biochim Biophys Acta, 1998, 1365:215~219.
[7] Newmeyer D, Farschon DM, Reed JC, et al. Cell-free apoptosis in Xenopus egg extracts: inhibition by Bcl-2 and requirement for an organelle fraction enriched in mitochondria[J]. Cell, 1994, 79:353.
, 百拇医药
[8] Li Y, Chopp M, Powers C, et al. Apoptosis and protein expression after focal cerebral ischemia in rat[J]. Brain Res, 1997, 765:301~312.
[9] Green DR, Reed JC. Mitochondria and apoptosis[J]. Science, 1998, 281:1309~1312.
[10] Li P. Cytochrome C and dATP-dependent formation of Apaf-f/caspase-9 complex initiates an apoptotic protease cascade[J]. Cell, 1997, 91:479.
[11] Fujimura M, Fujimura YM, Murakami K, et al. Cytosolic redistribution of cytochrome C after transient focal cerebral ischemia in rats[J]. J Cereb Blood Flow Metab, 1998, 18:1239~1247.
, http://www.100md.com
[12] Vander Heiden MG, Chandel NS, Williamson EK, et al. Bcl-xL regulates the membrane potential and volume homeostasis of mitochondria[J]. Cell, 1997, 91:627.
[13] Saikumar P, Zheng D, Weinberg JM, et al. Mechanisms of cell death in hypoxia/reoxygenation injury[J]. Oncogene, 1998, 17:3341~3349.
[14] Kluck RM, Bossy-Wetzel E, Green DR, et al. Prevention of apoptosis by Bcl-2: release of cytochrome C from mitochondria blocked[J]. Science, 1997, 275:1129.
, 百拇医药
[15] Walker JE. The NADH:ubiquinone oxidoreductase(complexⅠ) of respiratory chains[J]. Q Rev Biophys, 1992, 25:253~324.
[16] Budd SL. Mechanisms of neuronal damage in brain hypoxia/ischemia focus on the role of mitochondrial calcium accumulation[J]. Pharmacol Ther, 1998, 80:203~229.
[收稿日期] 1999-10-14 [修回日期] 2000-01-21, 百拇医药
单位:袁有忠(长海医院血液科,上海 200433);吴琦 邓小明 熊源长 刘树孝(上海第二军医大学附属长海医院麻醉科, 上海 200433)
关键词:局部缺血;再灌注损伤;基因表达;DNA,线粒体;大鼠
中国病理生理杂志000813
[摘 要] 目的:研究海马神经元缺血再灌注后线粒体相关基因的表达。方法:改良Pulsinell四血管阻断模型致大鼠脑缺血10min再灌注24 h后,取新鲜海马组织抽提mRNA。逆转录后经液相杂交递减无差异表达序列。以抑制性PCR方式扩增差异表达序列。之后构建cDNA文库,测序鉴定阳性克隆。最后应用斑点杂交确认差异表达。结果:构建cDNA文库后获得的78个阳性克隆中有7个克隆经最后的鉴定具有差异表达性。测序提示为细胞色素氧化酶C亚单位和NADH-泛醌氧化酶复合体4链的表达性片段。结论:细胞色素氧化酶C亚单位的差异表达从另一个侧面证实了之前提出的再灌注损伤中细胞色素氧化酶C的重要作用。再灌注损伤后NADH-泛醌氧化酶复合体4链的表达提示原来功能不明的NADH-泛醌氧化酶复合体β亚基可能在这一病理过程中发挥一定的作用。
, 百拇医药
[中图分类号] R743.31 [文献标识码] A
[文章编号] 1000-4718(2000)08-0722-04
Mitochondria respiratory chain related genes expression
in post ischemia/reperfusion rat
WU Qi, DENG Xiao-ming, XIONG Yuan-chang, LIU Shu-xiao
(Department of Anesthesiology, The Second Military Medical University Affiliated Changhai Hospital, Shanghai 200433,China)
, 百拇医药
YUAN You-zhong
(Department of Hematology, The Second Military Medical University Affiliated Changhai Hospital, Shanghai 200433,China)
[Abstrac] AIM: Apply the newly invented method of suppression subtractive hybridization to scan the mitochondria genes expression of hippocampus neurons in post ischemia/reperfusion rats. METHODS: Decapitate the rats suffering 10 min of whole brain ischemia and 24 h of reperfusion in Pulsinelli’s 4 vessels occlusion ischemia/reperfusion model. Dissociated hippocampus and isolated mRNA. After reversed transcription, use SSH to subtract the common sequences, then use suppressed PCR to amplify the differentially expressed sequences. After cloning and DNA sequencing, the dot hybridization was used to finally identify the differential gene expression. RESULTS: In the 78 positive clones acquired from cloning, 7 clones represent differential gene expression after confirmed by clone PCR, dot blot and DNA sequencing. The DNA sequencing data indicate these clones are ESTs of Cytochrome c oxidase subunit and NADH-ubiquinone oxidoreductase chain 4 gene. CONCLUSION: Cytochrome C oxidase mRNA level changs during brain ischemia /reperfusion. The differential expression of NADH-ubiquinone oxidoreductase chain 4, one of those 13 subunits of subcomplex β, suggests that subcomplex β may have important function associated with pathophysiologic changs in reperfusion injury.
, 百拇医药
[MeSH] Ischemia;Reperfusion injuries;Gene expression;DNA,mitochondrial;Rats
脑缺血/再灌注损伤后常会观察到原有的一些神经功能损害程度并不是静止在缺血的当时,而是进行性加重。这是因为调节这些功能的神经元发生延迟性死亡了。这种病理变化通常是凋亡。发生凋亡的神经元在受到致死性刺激的最初48 h内还能保持结构的完整。这提示:凋亡是一个过程,如果在这时间之内能有一些有效措施阻止凋亡的级联效应,那么挽救凋亡神经元是可能的。凋亡还是一种耗能的程序性死亡。线粒体不仅因为它是能量转换器而涉及到凋亡的调控中,更有证据表明线粒体的某些组份直接参与了凋亡的调控过程[1]。Diatchenko等[2]报道的抑制递减杂交法(suppression subtractive hybridization, SSH)能有效检测两个不同细胞群差异表达基因的方法。本研究应用该方法检测缺血再灌注后线粒体基因的表达。
, 百拇医药
材 料 和 方 法
一、脑缺血模型复制与分组
选用雄性SD大鼠,体重250~300 g。脑缺血模型采用改良的Pulsinelli法复制,即异戊巴比妥麻醉下烧闭双侧椎动脉同时双侧颈总动脉套线。24 h后,动物清醒下,拉紧双侧缝线并保持一定张力,阻断双侧颈总动脉造成脑缺血10 min。剪断缝线并抽出,开放颈总动脉血流造成再灌注24 h。正常对照组与脑缺血组动物各6只,断头处死大鼠,打开颅腔,取出海马组织后迅速放入液氮中速冻。24 h后转移至-85℃低温冰箱中保存。
二、海马组织mRNA的抽提、鉴定和浓缩
液氮冷冻条件下将海马组织捣碎成粉末。应用组织/细胞总RNA抽提试剂盒(Watson公司)抽提总RNA。随后应用mRNA Isolation Kit for Blood/Bone Marrow(Boehringer-mannheim,Germany)抽提mRNA,即polyA(+)RNA。
, 百拇医药
三、抑制性递减杂交分析
未经特别说明,试剂主要由PCR-SelectTM cDNA Subtraction kit(Clontech, USA)提供。
cDNA合成引物:5′-TTTTGTACAAGCTT30N1N-3′。
连接器1序列: 5′-CTAATACGACTCACTATAGGGCTCGAGCGGCCGCCCGGGCAGGT-3′
3′-GGCCCGTCCA-5′
连接器2序列: 5′-CTAATACGACTCACTATAGGGCAGCGTGGTCGCGGCCGAGGT-3′
3′-GCCGGCTCCA-5′
, 百拇医药
PCR引物1: 5′-CTAATACGACTCACTATAGGGCTC-3′
巢式PCR引物1: 5′-TCGAGCGGCCGCCCGGGCAGGT-3′
巢式PCR引物2: 5′-AGCGTGGTCGCGGCCGAGGT -3′
(一)mRNA逆转录合成cDNA 模板采用经分光光度计精确定量的mRNA (0.5 μg/μL)。
(二)限制性酶切反应及连接器的连接反应 cDNA产物纯化后经RsaⅠ限制性酶切后,实验组酶切产物分成相同的两组,分别接上连接器1和连接器2。
(三)液相杂交 第一轮杂交是由两组独立的杂交反应组成的, 杂交体系的构成为RsaⅠ限制性酶切后的对照组cDNA,带连接器的实验组cDNA(两组杂交反应分别含不同的连接器),1×杂交缓冲液。第一轮杂交的反应参数(98℃变性1.5 min, 68 ℃保温8 h)。第一轮杂交后进入第二轮杂交反应。在同一杂交体系中混合上述两组独立的杂交反应。同时在杂交体系中加入新鲜变性的RsaⅠ限制性酶切后的对照组cDNA,造成杂交体系中该成分过量。第二轮杂交的反应参数(68℃杂交过夜)。
, 百拇医药
(四)抑制PCR 第一轮PCR体系中,模板为第二轮杂交产物,PCR引物为PCR引物1。第二轮PCR体系中,模板为第一轮PCR产物,PCR引物为巢式PCR引物1和巢式PCR引物2。
四、cDNA文库的构建和测序分析
利用T/A cloning kit(Promega,USA),将第二轮PCR扩增产物插入T-Vector中。根据α 互补的原理,在涂布X-gal的琼脂平板上筛选出带插入片断的菌落。进一步以菌落PCR鉴定。抽提质粒进行全自动DNA测序。质粒的抽提采用Plasmid Mini Kit(Qiagen,USA)。 通过e-mail与genbank联系,借助BLAST程序分析核酸及蛋白的同源性。
五、斑点杂交
将送测序的质粒中的插入片断制备成[α-32P]dATP标记的探针,[α-32P]dATP购自北京亚辉生物医学工程公司,探针标记的试剂来自MegaprimeTM DNA labelling systems(Life science,USA)。将大约1 μg正常组和实验组总RNA直接烤膜固定到尼龙膜上(Hybrond, Germany)。杂交过程和洗膜过程参照《分子克隆实验指南》[3]。杂交膜在cyclone磷屏成像系统中曝光、扫描成像后储存于计算机中。
, 百拇医药
结 果
初步构建了大鼠脑缺血再灌注后差异表达的cDNA 文库。根据α互补的原理筛选出78个阳性克隆。同时由于差异表达cDNA 在构建文库前已被RsaⅠ限制性酶切,片断大小约为0.1~2 kb。 因而在菌落PCR中可借助来进一步鉴别阳性克隆。进一步筛选后得到12个阳性克隆(图1)。阳性克隆抽提质粒后行全自动测序分析。其中1、2、5、10、11、12号克隆对应细胞色素氧化酶C亚单位;3号克隆为ATP合成酶e链;4号克隆为NADH-泛醌氧化酶复合体4链(ND4);6号克隆为40S核糖体蛋白;7号克隆为硫糖蛋白-2;8号克隆为tRNA; 9号克隆为ATP合成酶a链。选择4、5、7、9号克隆,按材料方法中所述步骤进一步以斑点杂交验证差异表达性,结果4、5号克隆显示了差异表达(图2)。
NADH-泛醌氧化酶复合体4链的表达片段资料为:
3'TCACAGGAATATACTCAATATATATGATTATCACAAC
, 百拇医药
CCAACGAGGAAAACTAACCAGCCACATAAACAACCT
CCAACCTTCCCACACACGAGAATTAACACTCATAGCT
CTACACATTATTCCCCTCATACTATTAACAATCAACCC
TAAACTCATCACAGGCCTAACAATA 5'
细胞色素氧化酶C亚单位的的表达片段资料为:
3'TTGACTCCTACATAATCCCAACCAATGACCTAAAACC
AGGTGAACTTCGTCTATTAGAAGTTGATAATCGGGTA
GTCTTACCAATAGAACTTCCAATTCGTATACTAATCT
, http://www.100md.com
CATCCGAAGACGTCCTGCACTCATGAGCCATCCCTTC
ACTAGGGTTAAAAACCGACGCAATCCCCGGCCGCCTA
AACCAAGCTACTAGTCACATCAAACCGACCAGGTCTA
TTCTATGGCCAATGCTCTGAAATTTGCGGCTCAAATC
ACAGCTTCATACCCATTGTAC 5'
Fig 1 Positive clones selected by clone PCR
M: DNA marker(ΦX174DNA HaeⅢ digested fragment) ; lane 1~8,11,12~14 have almost the same size fragment, so they may represent one clone ; lane 9,10,15,16 have different size fragment, therefor probably represent different clone
, 百拇医药
图1 菌落PCR筛选出的阳性克隆
Fig 2 Dot hybridization of differential expressed clone
A: The 4th chain of NADH:ubiquinone oxidoreductase; B: Cytochrome C oxidase subunit; D: Control group; T: Test group
图2 差异表达克隆的斑点杂交分析
讨 论
试验采用的脑缺血模型是在Pulsinelli等创建的大鼠4条血管阻断所致全脑缺血模型的基础上进行的改进。我们曾观察该模型中阻断颈总动脉后局部脑血流量的变化,结果发现内侧中央前区、纹状皮层、小脑半球单小叶等脑区在脑缺血期间均显著下降,最低水平分别为缺血前的9%、11%、24%[4]。该模型造成的组织病理学变化、严重前脑缺血的重复性均良好,而且和经典Pulsinelli模型比较,阻断和开放颈总动脉系统更方便。
, http://www.100md.com
脑缺血再灌注后表现出差异表达的2个基因都是线粒体呼吸链的重要成分。 细胞色素氧化酶C(cytochrome C)位于线粒体内外膜间。它的功能是把电子从QH2-细胞色素氧化酶C还原酶复合体传递到细胞色素氧化酶C氧化酶复合体[5]。NADH-泛醌氧化酶复合体(NADH:ubiquinone oxidoreductase complex)也位于线粒体内外膜之间,它是电子进入呼吸链的第一道门户[6]。
线粒体长久以来是以能量转化中心的形象出现的,但最新的研究表明线粒体在细胞死亡的调控机制中发挥着重要的作用。在一项离体的细胞试验中发现,Bcl-2抑制核皱缩和DNA断裂的作用必须有线粒体参与[7]。但最早的突破是细胞色素氧化酶C注入培养细胞中引起凋亡表现的发现[8]。随后发现缺血再灌注损伤后细胞色素氧化酶C从线粒体中释放入胞浆中并和胱氨酸蛋白酶原-9(procaspase-9)和Apaf-1构成了“凋亡小体” ,进而激活其他半胱氨酸蛋白酶(caspase)共同完成一系列凋亡的生化过程[9,10]。还有学者观察到,大脑中动脉阻断90 min再灌注4 h, 细胞色素氧化酶C出现表达,再灌注24h达到高峰[11]。细胞色素氧化酶C的释放确切的机制不甚清楚。有研究提示线粒体内膜的通透性异常起了关键的作用。这种状态通常被定义为线粒体通透性转运(mitochondrial permeability transition, MPT),它允许分子量小于1500 D的分子自由扩散。当MPT成为不可逆变化后,线粒体内膜由于渗透压的作用极度皱缩,进而引起外膜破裂,细胞色素氧化酶C从内外膜间释放[12]。虽然线粒体外膜的破裂可以导致细胞色素氧化酶C、凋亡诱导因子(AIF)的释放,但是只有一部分线粒体的外膜破裂[13]。但有观察提示线粒体所有的细胞色素氧化酶C在凋亡时释放。这说明现有观察线粒体外膜破裂的手段不灵敏或者细胞色素氧化酶C的释放有其他机制[14]。
, 百拇医药
从牛心克隆到的NADH-泛醌氧化酶复合体由7个线粒体编码的亚基ND1- ND6(分子量50 kD左右)、ND4L和34个核编码的亚单位构成。对NADH-泛醌氧化酶复合体的结构功能分析显示:该复合体由 α、β亚单位构成;α亚单位含28个亚基,包括了全部的黄素蛋白和铁蛋白,集中了全部的具有还原活性的Fe-S簇,是NADH-泛醌氧化酶复合体氧化还原活性中心所在;β亚单位由13个亚基构成(包含ND4), 无氧化还原活性中心,其生化功能不明[15]。在NADH-泛醌氧化酶复合体中存在如此庞大、复杂的但却与主要功能无关的β亚单位,人们不禁怀疑它是否具有其他重要的生理功能?有学者提出凋亡调控过程中,MPT后线粒体外膜破裂,凋亡诱导因子(一类分子量50 kD左右的分子)从内外膜间释放,凋亡的下游过程启动[16]。NADH-泛醌氧化酶复合体4链(ND4)的分子量、定位、缺血再灌注后表达等特征,使之类似于凋亡诱导因子。但ND4和细胞死亡调控的确切关系需要进一步的严谨实验证实。
, 百拇医药
[参 考 文 献]
[1] Abe K, Aoki M, Kawagoe J, et al. Ischemic delayed neuronal death: A mitochondrial hypothesis[J]. Stroke, 1995, 26:1473~1489.
[2] Diatchenko L, Lau YFC, Campbell AP, et al. Suppression subtractive hybridization: A method for generating differentially regulated or tissue-specific cDNA probes and libraries[J]. Proc Natl Acad Sci, 1996, 93:6025~6030.
[3] J 萨姆布鲁克,EF弗里奇,T曼尼阿蒂斯著.分子克隆实验指南[M].第2版.北京:科学出版社,1996.65~68.
, http://www.100md.com
[4] 邓小明,石一鸣,王景阳,等.脑缺血再灌注损伤的动物模型[J].中国病理生理杂志, 1991, 7:208.
[5] Boyer PD, Chance B, Ernster L, et al. Oxidative phosphorylation and photo-phosphorylation[J]. Annu Rev Biochem, 1977, 46:955~1026.
[6] Friedrich T, Abelmann A, Brors B, et al. Redox components and structure of the respiratory NNDH:ubiquinone oxidoreductase(complexⅠ)[J]. Biochim Biophys Acta, 1998, 1365:215~219.
[7] Newmeyer D, Farschon DM, Reed JC, et al. Cell-free apoptosis in Xenopus egg extracts: inhibition by Bcl-2 and requirement for an organelle fraction enriched in mitochondria[J]. Cell, 1994, 79:353.
, 百拇医药
[8] Li Y, Chopp M, Powers C, et al. Apoptosis and protein expression after focal cerebral ischemia in rat[J]. Brain Res, 1997, 765:301~312.
[9] Green DR, Reed JC. Mitochondria and apoptosis[J]. Science, 1998, 281:1309~1312.
[10] Li P. Cytochrome C and dATP-dependent formation of Apaf-f/caspase-9 complex initiates an apoptotic protease cascade[J]. Cell, 1997, 91:479.
[11] Fujimura M, Fujimura YM, Murakami K, et al. Cytosolic redistribution of cytochrome C after transient focal cerebral ischemia in rats[J]. J Cereb Blood Flow Metab, 1998, 18:1239~1247.
, http://www.100md.com
[12] Vander Heiden MG, Chandel NS, Williamson EK, et al. Bcl-xL regulates the membrane potential and volume homeostasis of mitochondria[J]. Cell, 1997, 91:627.
[13] Saikumar P, Zheng D, Weinberg JM, et al. Mechanisms of cell death in hypoxia/reoxygenation injury[J]. Oncogene, 1998, 17:3341~3349.
[14] Kluck RM, Bossy-Wetzel E, Green DR, et al. Prevention of apoptosis by Bcl-2: release of cytochrome C from mitochondria blocked[J]. Science, 1997, 275:1129.
, 百拇医药
[15] Walker JE. The NADH:ubiquinone oxidoreductase(complexⅠ) of respiratory chains[J]. Q Rev Biophys, 1992, 25:253~324.
[16] Budd SL. Mechanisms of neuronal damage in brain hypoxia/ischemia focus on the role of mitochondrial calcium accumulation[J]. Pharmacol Ther, 1998, 80:203~229.
[收稿日期] 1999-10-14 [修回日期] 2000-01-21, 百拇医药