牛泡沫病毒内部启动子的克隆及功能分析
作者:张莉 王世珍 刘佳建 刘淑红 陈启民 耿运琪
单位:南开大学生命科学学院,天津,300071
关键词:牛泡沫病毒;env基因;内部启动子;LTR
病毒学报000308 摘要:以该实验室分离并鉴定的牛泡沫病毒(BFV)中国毒株(BFV3026)为材料,用PCR方法首次克隆了位于env基因3′端的内部启动子,经序列分析后,引入luc基因作瞬时表达分析。结果表明,该内部启动子不但基础活性高于LTR,而且转录活性在Tas参与下被大大激活,其激活活性也远远高于LTR。同源分析表明,非灵长类泡沫病毒内部启动子之间的同源性高于其与灵长类泡沫病毒内部启动子之间的同源性。
中图分类号:S852.6+9.3; Q75 文献标识码:A
, 百拇医药
文章编号:1000-8721(2000)03-0227-05
Cloning and Functional Analysis of the Internal Promoter of a Bovine Foamy Virus
ZHANG Li,WANG Shi-zhen,LIU Jia-jian,LIU Shu-hong,CHEN Qi-min,GENG Yun-qi
(College of Life Sciences, Nankai University, Tianjin 300071, China)
Abstract: Bovine foamy virus is a member of spumavirus that encodes three retroviral genes, gag, pol and env for virion proteins as well as additional open reading frames. ORF-1 was proven to be a viral transactivator which augmented transcription directed by the long terminal repeat (LTR) through cis-acting targets in the U3 domain of the LTR. Recently, we demonstrated by using sequence analysis and transient expression assay that a newly isolated BFV3026 strain also encoded an internal promoter (IP) in the 3' end of BFV env gene. Transcription directed by the IP of BFV was greatly activated by the ORF-1 in BL-12 cell line. Furthermore, both the basal activity and enhanced activity of IP were much greater than those of LTR, which suggests that different transactivation mechanisms were involved in regulating BFV LTR and IP.
, 百拇医药
Key words: Bovine foamy virus; env gene; internal promoter; long terminal repeat (LTR)
泡沫病毒(spumaviruses)在分类上属反转录病毒科(Retroviridae)的泡沫病毒属(Spumavirus)。泡沫病毒感染细胞后形成多核大细胞,即合胞体,并诱发细胞质形成空泡和内质网水肿,从而使被感染的细胞看似泡沫状,泡沫病毒因此得名。目前已从健康或患病的人、猴、猫、牛等体内分离到人泡沫病毒(human foamy virus,HFV)、猴泡沫病毒1、3型(simian foamy virus type 1、3,SFV-1、SFV-3)、猫泡沫病毒(feline foamy virus,FeFV)、牛泡沫病毒(bovine foamy virus,BFV)等。到目前为止有关泡沫病毒的研究大部分集中在HFV、SFV等灵长类泡沫病毒上,而对于非灵长类泡沫病毒,特别是BFV的研究还很不深入。BFV全序列近期才见报道[1],其基因组全长12.0kb,两端为1.30kb的长末端重复序列(LTR),中间部分除gag、pol、env三个结构基因外,在env与3′LTR之间有两个重叠的开放阅读框架ORF-1和ORF-2,编码自身的反式激活因子(transactivator of spumaviruses,Tas)。Tas不仅对自身LTR行使功能,而且能激活慢病毒的基因表达。近年来在泡沫病毒家族成员HFV[2,3]、SFV-1型[4]、SFV-3型[5]中陆续发现了除5′LTR之外的第二类启动子。该启动子位于env基因内部,调节蛋白Tas编码区近上游,称为内部启动子(internal promoter,IP)。该启动子亦依赖于Tas。在牛泡沫病毒中是否存在内部启动子尚未见报道。
, 百拇医药
材料与方法
1 病毒培养与细胞 BFV3026毒株是本室从BIV检测为阳性的3026号牛外周血中分离得到的,经分子生物学鉴定,证明其为一株新的泡沫病毒[6]。新生牛肺细胞(newborn bovine lung cells,NBL)用于BFV3026病毒株的繁殖。牛肺细胞系BL-12用于转染。
2 病毒Hirt DNA的提取 反转录病毒感染宿主细胞后首先反转录成双链DNA,在反转录病毒感染的细胞的细胞核内常常存在一些非整合的线形或环状双链DNA分子,通常称为Hirt DNA。按文献的方法从被BFV3026病毒株感染的NBL细胞中抽提NBL/BFV3026的Hirt DNA,并用TE缓冲液溶解。
3 引物的设计与合成 有关泡沫病毒内部启动子的序列尚无报道,本文根据BFV基因组序列,参与HFV、SFV-1的IP保守序列,化学合成1对引物P8564和P9488,旨在通过PCR扩增完整的BFV内部启动子。引物序列如下: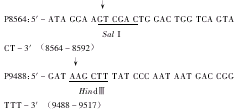
, 百拇医药
4 PCR反应 以NBL/BFV3026 Hirt DNA为模板进行PCR反应。反应条件为94℃4min,55℃1min,72℃3min,1个循环;94℃30s,55℃ 1min,72℃3min,35个循环;94℃30s,55℃1min,72℃10min,1个循环。扩增产物于1%琼脂糖凝胶电泳,观察照相。
5 克隆与测序 将BFV3026的PCR产物直接克隆到pGEM-T载体上(参见Promega产品说明书),并用SP6和T7引物对其进行序列分析。
6 序列分析 核苷酸的序列对比与分析采用Dnasis计算机分析软件(Hitachi Software Engineering Co. Ltd.)。
7 功能质粒的构建 以PCR方法扩增得到954bp内部启动子片段,将其正向连至载体pGEM-T上得到pT-BFVIP+(8564/9517)(括号内数字为基因组编号)。经酶切鉴定正确之后,将其上的BFV IP片段亚克隆到质粒pBFVLTR-luc(本室保存)上替换BFV LTR 区段得到了报告质粒pBFVIP(8572/9509)-luc。该质粒的报告基因luciferase上游连有BFV完整的内部启动子。pRSV-BORF-1激活质粒由本室构建,即BFV反式激活因子Tas(ORF-1)基因连在RSV启动子下游。
, http://www.100md.com
8 体外瞬时表达实验 牛肺细胞系BL-12以3×105ml-1接种于6孔板,每孔2ml,于37℃ 5%CO2条件下,10%胎牛血清DMEM培养基中培养。转染采用阳离子脂质体法(LipofectAMINE Reagent)。具体转染步骤参见GIBCO/BRL产品说明书。2天后裂解细胞测定Luc酶活力。细胞裂解液和Luc酶活力测定试剂盒购自Promega公司。按试剂盒提供的方法,用生物化学发光测量仪(上海检测技术所检测仪器厂生产)测定化学发光强度,方法原理见文献[7],Luc酶的活力以4S内的光强度表示。
结 果
1 完整的牛泡沫病毒内部启动子的克隆与序列分析
本文根据文献报道[1]选择泡沫病毒中env基因3′端,相对保守的基因区段设计并合成BFV8564、BFV9488引物,以NBL/BFV3026 Hirt DNA为模板进行PCR,得到954 bp特异性带。而以NBL细胞DNA为模板或体系中不加任何DNA时则没有任何产物。回收954bp特异性PCR产物,直接与pGEM-T载体连接,转化E.coli JM103。使用IPTG,X-gal筛选白色转化子,得到质粒pT-BFVIP(8564/9517)。
, http://www.100md.com
采用SP6和T7引物,对克隆的8564~9517基因组区段进行序列分析,并将结果与已报道的BFV美国毒株基因组进行比较,发现在这954bp的片段中发生了7个碱基的改变。分别是8 583位G变成A,8 598位A变成G,8 722位A变成G,8 778位A变成G,9 061位C变成A,9 185位C变成T,9 361位G变成A,序列相似度达到99.3%。该片段中含有典型的真核启动子TATA盒(图1)。根据HFV、SFV-1中已有的报道,推测这段序列中含有完整的内部启动子(IP)。
2 牛泡沫病毒内部启动子功能分析
2.1 功能质粒构建 为了测定IP的启动子活性,我们构建了IP的报告质粒。将质粒pT-BFVIP(8564/9517)用SalⅠ和HindⅢ酶切后,回收938bp小片段。同样用SalⅠ和HindⅢ切pBFVLTR-luc质粒,回收5.82kb大片段。将上述两个回收片段连接后转化E.coli TG1经酶切鉴定得到质粒pBFVIP(8572/9509)-luc。该片段在荧火虫荧光素酶报告基因上游连有牛泡沫病毒完整的内部启动子。该片段构建过程见图2。
, 百拇医药
图1 克隆的牛泡沫病毒基因组8 564~9 517区段序列
Figure 1 The sequence of BFV genome 8 564~9 517
图2 pBFVIP(8572/9509)-luc报告质粒的构建过程
Figure 2 Construction of the reported plasmid of pBFVIP(8572/9509)-luc
2.2 功能分析 为测定内部启动子的活性,将含完整内部启动子的报告质粒pBFVIP(8 572/9 509)-luc单独转染BL-12,测定IP的基础活性;将pBFVIP(8 572/9 509)-luc与含有BFV Tas基因的激活质粒pRSV-BORF-1共转染牛肺细胞系BL-12,测定IP被激活后的活性及激活倍数。转染结果如表1。
, http://www.100md.com
表1 BFV LTR和IP的基础活性和激活活性比较
Table 1 Basal activity and augmented activity of BFV LTR and IP Reported
plasmid
Basal
activity
Augmented
activity
Multiple of
activation
Ratio of effector
, 百拇医药
plasmid and reported
plasmid at
maximum activation
pBFVIP-luc
175
25375
145
2.5:1
pBFVLTR-luc
15
315
21
, http://www.100md.com
1:1
β-gal normalizing system is introduced into each transfection. Doses of plasmids are showed in μg for convenience. Since sizes of pBFVLTR-luc and pBFVIP-luc are similar and both limited to 6.8~7.1kb,similar doses in μg can be approximately regarded as similar moles.0.2μg pBFVLTR-luc and pBFVIP-luc are used in transfection.
由表1可以看出:当pBFVIP与pRSV-BORF-1共同转染时,转录活性比本底提高了145倍,从而证明本文克隆的基因组片段8 572~9 509确实为牛泡沫病毒的内部启动子。为了比较该内部启动子与LTR的活性,本文亦将含有完整BFV LTR的报告质粒pBFVLTR(-55/1257)-luc单独转染或与pRSV-BORF-1共转染BL-12。相同用量下的pBFVIP-luc比pBFVLTR-luc转染细胞所得的Luc活性高得多,说明BFV IP的基础活性远远高于BFV LTR。当上述两个质粒分别与pRSV-BORF-1共转染细胞时,ORF-1对-55/1257全长LTR片段的最高激活倍数为21倍,出现在ORF-1表达质粒与报告质粒的用量比为1:1的情况下;而ORF-1对8 572/9 509全长IP片段的最高激活倍数达145倍,出现在两质粒用量比为2.5:1的情况下。这说明BFV IP不但也可被BFV反式激活因子ORF-1显著激活,而且激活倍数远远高于ORF-1对BFV LTR的激活。此外,IP与LTR达到最大激活倍数所需ORF-1用量的差异也暗示了ORF-1可能通过不同的机制激活IP与LTR。
, 百拇医药
3 不同病毒内部启动子同源性比较
将克隆的BFV内部启动子分别与HFV、SFV-1、SFV-3、FeFV env基因相应区段进行比较分析同源性。其它四种病毒的比较区段分别为HFV:8 847~9 347;SFV-1:8 011~8 511,SFV-3:9 362~9 862,FeFV:8 587~9 097。TATA盒均用含阴影的方格标出来。5种病毒中相同的碱基用星号标出。H、S表示HFV、SFV内部启动子转录本起始5′端(图3)。

图3 HFV、SFV-1、SFV-3、BFV、FeFV内部启动子及附近区域序列比较
Figure 3 Sequence alignment of the region of env encompassing the internal promoters of HFV,SFV-1,SFV-3,BFV,FeFV
, http://www.100md.com
Asterisks(*) show 100% identical nucleotides in the five foamy viruses.Shaded frame marks their TATA boxes.
序列同源性分析表明,BFV与其它几种病毒同源程度分别为:BFV与SFV-1为44.1%,BFV与SFV-3为44.5%;BFV与HFV为46.5%,BFV与FeFV为61.4%。而SFV-1与SFV-3有77%同源,SFV-3与HFV有69%同源,SFV-1与HFV有66%同源。由此可见:灵长类泡沫病毒之间,尤其是相同种属的泡沫病毒之间,如SFV-1、SFV-3同源度较高;非灵长类泡沫病毒,如BFV和FeFV之间同源度也比较高。而灵长类泡沫病毒与非灵长类泡沫病毒之间同源度相对较低。具体数据见表2。
表2 牛泡沫病毒内部启动子与其它泡沫病毒内部启动子同源性比较(%)
Table 2 Pairwise homologies for the IP sequence of BFV and other foamy viruses (%) BFV-IP
, http://www.100md.com
8914-9413
9048-9413
9112-9413
9200-9413
BFV-FeFV
61.4
57.9
58.9
61.7
BFV-HFV
46.5
48.1
, 百拇医药
54.3
61.9
BFV-SFV1
44.1
46.4
52.0
58.1
BFV-SFV3
44.5
48.6
55.6
66.4
讨 论
, 百拇医药
本研究从本实验室分离并鉴定的BFV3026中国毒株中克隆到了完整的内部启动子,并构建了pBFVIP-luc表达质粒。体外瞬时表达实验表明,该启动子可以被牛泡沫病毒自身编码的反式激活因子ORF-1所激活,从而证实了牛泡沫病毒中内部启动子的存在。IP的存在使泡沫病毒的基因表达调节机制更显复杂。以前的一些研究表明:HFV的env IP区域为8 921~9 332,TATA盒位于转录起始点上游的9 164~9 169,SFV-1 env IP位于8 183~8 370,其Tas调控位点位于8 249~8 370,TATA盒为8 380~8 385。SFV3 env IP区域为9 534~9 820,9 731~9 736为其TATA盒。而本文克隆的BFV env 3′区段(基因组8 572~9 509)中包含了完整的牛泡沫病毒内部启动子。该启动子TATA盒位于基因组9 282~9 285之间。泡沫病毒的TATA盒序列及附近序列十分保守,均为TATAAAAGA*CA*AT。该TATA盒很可能与TATA盒结合因子TFIID相互作用,而该因子是宿主细胞RNA聚合酶Ⅱ复合物起始转录所必需的。核苷酸序列同源分析表明,BFV env基因3′端与HFV、SFV-1、SFV-3、FeFV同源性均达44%以上。其中与FeFV的同源性最高,达61.4%。但当区段逐渐向TATA盒靠近时,同源性逐步升高(表1)。这暗示着越靠近TATA盒序列保守性越强,很可能在TATA盒近上游包含有重要的调控位点或ORF-1反式激活的靶位点(TREIP)。但仅根据同源性分析还难以作出定论,而有待于对该区段进一步缺失或定点突变来确定TREIP的位置。
, 百拇医药
BFV基因组中存在两个启动子的意义有待于进一步阐明,与其它病毒相比,泡沫病毒中内部启动子的存在并非独特,鼠乳腺肿瘤病毒(mouse mammary tumor viruses)和人T细胞白血病病毒1型(human T-cell leukemia virus type 1)亦可以从LTR及env基因内部起始转录,但这些病毒的内部启动子不能被相应病毒编码的某种蛋白反式激活。而BFV IP及LTR均强列依赖牛泡沫病毒自身编码的反式激活因子ORF-1。尽管这两种启动子均能被ORF-1反式激活,但调控机制却不尽相同。将BFV IP TATA附近区域与LTR相应区段,以及BFV IP TATA盒上游区域与LTR U3区内的两个TRELTR进行比较(TREI:-983/-668,TREII:-470/-140,转录起始位点为+1)[8],均只发现极短簇的同源区。IP和LTR较低的同源性,暗示着两个启动子的反式激活可能有不同的因子参与。Mergia在对SFV-1的研究中曾提出过“时序模式(temporal pattern)”[5]。该模式认为,和许多DNA病毒一样,泡沫病毒在复制周期的不同时期,由不同的阶段特异性启动子(stage-specific promoter)控制不同的基因表达。即表达受2个或多个启动子调控。在病毒感染的早期,首先由IP起始转录,表达出病毒的调节蛋白Tas;随后,Tas一方面作用于LTR,表达出病毒结构蛋白,完成病毒颗粒的组装;另一方面反馈激活IP,合成更多的调节蛋白。由于至今还没有在泡沫病毒中发现与Rev/Rex相当的转录后调节蛋白,所以IP的存在在功能上替代了其它反转录病毒的转录后调节蛋白的作用。本文瞬时表达分析的结果也正好符合该模式,无BFV ORF-1存在时,即相当于病毒感染的早期,BFV IP的基础活性远远高于LTR,能够引发少量调节蛋白的表达;而有ORF-1存在时,即相当于病毒凭借IP的基础活性表达出部分调节蛋白后,IP的活性和LTR的活性均被大大激活。这一结果支持了“时序模式”,揭示了BFV与HFV、SFV-1、SFV-3有着相似的基因表达调控机制。
, 百拇医药
基金项目:国家自然科学基金(39770032);教育部博士点基金资助项目(98005516)
作者简介:张莉(1973-),女,湖南人,分子遗传学硕士。
联系作者:耿运琪
参考文献:
[1] Holzschu D L, Delaney M A, Renshaw R W, et al. The nucleotide sequence and spliced pol mRNA levels of the nonprimate spumavirus, bovine foamy virus [J] . J Virol, 1998, 72 (3) : 2177-2182.
[2] Lochelt M, Flugel R M, Aaboud M, et al. Human foamy virus genome possesses an internal, Bel-1-dependent and functional promoter [J] . Proc Natl Acad Sci USA, 1993, 90: 7317-7321.
, 百拇医药
[3] Lochelt M, Muranyi W, Flugel R M, et al. The human foamy virus internal promoter directs the expression of the functional bel-1 transactivator and bet protein early after infection [J] . J Virol, 1994, 68 (20) : 638-645.
[4] Mergia A. Simian foamy virus type 1 contains a second promoter located at the 3' end of the env gene [J] . Virology, 1994, 199: 219-222
[5] Campbell M, Renshaw G L, Renne R, et al. Characterization of the internal promoter of simian foamy viruses [J] . J Virol, 1994, 68 (8) : 4811-4820
[6] 刘淑红,陈荷新,陈家童,等. 牛泡沫病毒(BSV)3026毒株的分离及分子生物学鉴定[J].病毒学报,1997,13(2):140-145.
[7] 刘国文,纪永刚,梁臣,等.萤火虫荧光素酶基因构建BIV-LTR启动子表达体系[J].病毒学报,1994,10(4):377-380.
[8] 刘佳建,王世珍,张莉,等.牛泡沫病毒调节蛋白功能及其在LTR上应答元件的研究[J].病毒学报,2000,16(2):141~149.
来稿日期:1999-02-01;修回日期:1999-11-29, http://www.100md.com
单位:南开大学生命科学学院,天津,300071
关键词:牛泡沫病毒;env基因;内部启动子;LTR
病毒学报000308 摘要:以该实验室分离并鉴定的牛泡沫病毒(BFV)中国毒株(BFV3026)为材料,用PCR方法首次克隆了位于env基因3′端的内部启动子,经序列分析后,引入luc基因作瞬时表达分析。结果表明,该内部启动子不但基础活性高于LTR,而且转录活性在Tas参与下被大大激活,其激活活性也远远高于LTR。同源分析表明,非灵长类泡沫病毒内部启动子之间的同源性高于其与灵长类泡沫病毒内部启动子之间的同源性。
中图分类号:S852.6+9.3; Q75 文献标识码:A
, 百拇医药
文章编号:1000-8721(2000)03-0227-05
Cloning and Functional Analysis of the Internal Promoter of a Bovine Foamy Virus
ZHANG Li,WANG Shi-zhen,LIU Jia-jian,LIU Shu-hong,CHEN Qi-min,GENG Yun-qi
(College of Life Sciences, Nankai University, Tianjin 300071, China)
Abstract: Bovine foamy virus is a member of spumavirus that encodes three retroviral genes, gag, pol and env for virion proteins as well as additional open reading frames. ORF-1 was proven to be a viral transactivator which augmented transcription directed by the long terminal repeat (LTR) through cis-acting targets in the U3 domain of the LTR. Recently, we demonstrated by using sequence analysis and transient expression assay that a newly isolated BFV3026 strain also encoded an internal promoter (IP) in the 3' end of BFV env gene. Transcription directed by the IP of BFV was greatly activated by the ORF-1 in BL-12 cell line. Furthermore, both the basal activity and enhanced activity of IP were much greater than those of LTR, which suggests that different transactivation mechanisms were involved in regulating BFV LTR and IP.
, 百拇医药
Key words: Bovine foamy virus; env gene; internal promoter; long terminal repeat (LTR)
泡沫病毒(spumaviruses)在分类上属反转录病毒科(Retroviridae)的泡沫病毒属(Spumavirus)。泡沫病毒感染细胞后形成多核大细胞,即合胞体,并诱发细胞质形成空泡和内质网水肿,从而使被感染的细胞看似泡沫状,泡沫病毒因此得名。目前已从健康或患病的人、猴、猫、牛等体内分离到人泡沫病毒(human foamy virus,HFV)、猴泡沫病毒1、3型(simian foamy virus type 1、3,SFV-1、SFV-3)、猫泡沫病毒(feline foamy virus,FeFV)、牛泡沫病毒(bovine foamy virus,BFV)等。到目前为止有关泡沫病毒的研究大部分集中在HFV、SFV等灵长类泡沫病毒上,而对于非灵长类泡沫病毒,特别是BFV的研究还很不深入。BFV全序列近期才见报道[1],其基因组全长12.0kb,两端为1.30kb的长末端重复序列(LTR),中间部分除gag、pol、env三个结构基因外,在env与3′LTR之间有两个重叠的开放阅读框架ORF-1和ORF-2,编码自身的反式激活因子(transactivator of spumaviruses,Tas)。Tas不仅对自身LTR行使功能,而且能激活慢病毒的基因表达。近年来在泡沫病毒家族成员HFV[2,3]、SFV-1型[4]、SFV-3型[5]中陆续发现了除5′LTR之外的第二类启动子。该启动子位于env基因内部,调节蛋白Tas编码区近上游,称为内部启动子(internal promoter,IP)。该启动子亦依赖于Tas。在牛泡沫病毒中是否存在内部启动子尚未见报道。
, 百拇医药
材料与方法
1 病毒培养与细胞 BFV3026毒株是本室从BIV检测为阳性的3026号牛外周血中分离得到的,经分子生物学鉴定,证明其为一株新的泡沫病毒[6]。新生牛肺细胞(newborn bovine lung cells,NBL)用于BFV3026病毒株的繁殖。牛肺细胞系BL-12用于转染。
2 病毒Hirt DNA的提取 反转录病毒感染宿主细胞后首先反转录成双链DNA,在反转录病毒感染的细胞的细胞核内常常存在一些非整合的线形或环状双链DNA分子,通常称为Hirt DNA。按文献的方法从被BFV3026病毒株感染的NBL细胞中抽提NBL/BFV3026的Hirt DNA,并用TE缓冲液溶解。
3 引物的设计与合成 有关泡沫病毒内部启动子的序列尚无报道,本文根据BFV基因组序列,参与HFV、SFV-1的IP保守序列,化学合成1对引物P8564和P9488,旨在通过PCR扩增完整的BFV内部启动子。引物序列如下:
, 百拇医药
4 PCR反应 以NBL/BFV3026 Hirt DNA为模板进行PCR反应。反应条件为94℃4min,55℃1min,72℃3min,1个循环;94℃30s,55℃ 1min,72℃3min,35个循环;94℃30s,55℃1min,72℃10min,1个循环。扩增产物于1%琼脂糖凝胶电泳,观察照相。
5 克隆与测序 将BFV3026的PCR产物直接克隆到pGEM-T载体上(参见Promega产品说明书),并用SP6和T7引物对其进行序列分析。
6 序列分析 核苷酸的序列对比与分析采用Dnasis计算机分析软件(Hitachi Software Engineering Co. Ltd.)。
7 功能质粒的构建 以PCR方法扩增得到954bp内部启动子片段,将其正向连至载体pGEM-T上得到pT-BFVIP+(8564/9517)(括号内数字为基因组编号)。经酶切鉴定正确之后,将其上的BFV IP片段亚克隆到质粒pBFVLTR-luc(本室保存)上替换BFV LTR 区段得到了报告质粒pBFVIP(8572/9509)-luc。该质粒的报告基因luciferase上游连有BFV完整的内部启动子。pRSV-BORF-1激活质粒由本室构建,即BFV反式激活因子Tas(ORF-1)基因连在RSV启动子下游。
, http://www.100md.com
8 体外瞬时表达实验 牛肺细胞系BL-12以3×105ml-1接种于6孔板,每孔2ml,于37℃ 5%CO2条件下,10%胎牛血清DMEM培养基中培养。转染采用阳离子脂质体法(LipofectAMINE Reagent)。具体转染步骤参见GIBCO/BRL产品说明书。2天后裂解细胞测定Luc酶活力。细胞裂解液和Luc酶活力测定试剂盒购自Promega公司。按试剂盒提供的方法,用生物化学发光测量仪(上海检测技术所检测仪器厂生产)测定化学发光强度,方法原理见文献[7],Luc酶的活力以4S内的光强度表示。
结 果
1 完整的牛泡沫病毒内部启动子的克隆与序列分析
本文根据文献报道[1]选择泡沫病毒中env基因3′端,相对保守的基因区段设计并合成BFV8564、BFV9488引物,以NBL/BFV3026 Hirt DNA为模板进行PCR,得到954 bp特异性带。而以NBL细胞DNA为模板或体系中不加任何DNA时则没有任何产物。回收954bp特异性PCR产物,直接与pGEM-T载体连接,转化E.coli JM103。使用IPTG,X-gal筛选白色转化子,得到质粒pT-BFVIP(8564/9517)。
, http://www.100md.com
采用SP6和T7引物,对克隆的8564~9517基因组区段进行序列分析,并将结果与已报道的BFV美国毒株基因组进行比较,发现在这954bp的片段中发生了7个碱基的改变。分别是8 583位G变成A,8 598位A变成G,8 722位A变成G,8 778位A变成G,9 061位C变成A,9 185位C变成T,9 361位G变成A,序列相似度达到99.3%。该片段中含有典型的真核启动子TATA盒(图1)。根据HFV、SFV-1中已有的报道,推测这段序列中含有完整的内部启动子(IP)。
2 牛泡沫病毒内部启动子功能分析
2.1 功能质粒构建 为了测定IP的启动子活性,我们构建了IP的报告质粒。将质粒pT-BFVIP(8564/9517)用SalⅠ和HindⅢ酶切后,回收938bp小片段。同样用SalⅠ和HindⅢ切pBFVLTR-luc质粒,回收5.82kb大片段。将上述两个回收片段连接后转化E.coli TG1经酶切鉴定得到质粒pBFVIP(8572/9509)-luc。该片段在荧火虫荧光素酶报告基因上游连有牛泡沫病毒完整的内部启动子。该片段构建过程见图2。
, 百拇医药
图1 克隆的牛泡沫病毒基因组8 564~9 517区段序列
Figure 1 The sequence of BFV genome 8 564~9 517
图2 pBFVIP(8572/9509)-luc报告质粒的构建过程
Figure 2 Construction of the reported plasmid of pBFVIP(8572/9509)-luc
2.2 功能分析 为测定内部启动子的活性,将含完整内部启动子的报告质粒pBFVIP(8 572/9 509)-luc单独转染BL-12,测定IP的基础活性;将pBFVIP(8 572/9 509)-luc与含有BFV Tas基因的激活质粒pRSV-BORF-1共转染牛肺细胞系BL-12,测定IP被激活后的活性及激活倍数。转染结果如表1。
, http://www.100md.com
表1 BFV LTR和IP的基础活性和激活活性比较
Table 1 Basal activity and augmented activity of BFV LTR and IP Reported
plasmid
Basal
activity
Augmented
activity
Multiple of
activation
Ratio of effector
, 百拇医药
plasmid and reported
plasmid at
maximum activation
pBFVIP-luc
175
25375
145
2.5:1
pBFVLTR-luc
15
315
21
, http://www.100md.com
1:1
β-gal normalizing system is introduced into each transfection. Doses of plasmids are showed in μg for convenience. Since sizes of pBFVLTR-luc and pBFVIP-luc are similar and both limited to 6.8~7.1kb,similar doses in μg can be approximately regarded as similar moles.0.2μg pBFVLTR-luc and pBFVIP-luc are used in transfection.
由表1可以看出:当pBFVIP与pRSV-BORF-1共同转染时,转录活性比本底提高了145倍,从而证明本文克隆的基因组片段8 572~9 509确实为牛泡沫病毒的内部启动子。为了比较该内部启动子与LTR的活性,本文亦将含有完整BFV LTR的报告质粒pBFVLTR(-55/1257)-luc单独转染或与pRSV-BORF-1共转染BL-12。相同用量下的pBFVIP-luc比pBFVLTR-luc转染细胞所得的Luc活性高得多,说明BFV IP的基础活性远远高于BFV LTR。当上述两个质粒分别与pRSV-BORF-1共转染细胞时,ORF-1对-55/1257全长LTR片段的最高激活倍数为21倍,出现在ORF-1表达质粒与报告质粒的用量比为1:1的情况下;而ORF-1对8 572/9 509全长IP片段的最高激活倍数达145倍,出现在两质粒用量比为2.5:1的情况下。这说明BFV IP不但也可被BFV反式激活因子ORF-1显著激活,而且激活倍数远远高于ORF-1对BFV LTR的激活。此外,IP与LTR达到最大激活倍数所需ORF-1用量的差异也暗示了ORF-1可能通过不同的机制激活IP与LTR。
, 百拇医药
3 不同病毒内部启动子同源性比较
将克隆的BFV内部启动子分别与HFV、SFV-1、SFV-3、FeFV env基因相应区段进行比较分析同源性。其它四种病毒的比较区段分别为HFV:8 847~9 347;SFV-1:8 011~8 511,SFV-3:9 362~9 862,FeFV:8 587~9 097。TATA盒均用含阴影的方格标出来。5种病毒中相同的碱基用星号标出。H、S表示HFV、SFV内部启动子转录本起始5′端(图3)。
图3 HFV、SFV-1、SFV-3、BFV、FeFV内部启动子及附近区域序列比较
Figure 3 Sequence alignment of the region of env encompassing the internal promoters of HFV,SFV-1,SFV-3,BFV,FeFV
, http://www.100md.com
Asterisks(*) show 100% identical nucleotides in the five foamy viruses.Shaded frame marks their TATA boxes.
序列同源性分析表明,BFV与其它几种病毒同源程度分别为:BFV与SFV-1为44.1%,BFV与SFV-3为44.5%;BFV与HFV为46.5%,BFV与FeFV为61.4%。而SFV-1与SFV-3有77%同源,SFV-3与HFV有69%同源,SFV-1与HFV有66%同源。由此可见:灵长类泡沫病毒之间,尤其是相同种属的泡沫病毒之间,如SFV-1、SFV-3同源度较高;非灵长类泡沫病毒,如BFV和FeFV之间同源度也比较高。而灵长类泡沫病毒与非灵长类泡沫病毒之间同源度相对较低。具体数据见表2。
表2 牛泡沫病毒内部启动子与其它泡沫病毒内部启动子同源性比较(%)
Table 2 Pairwise homologies for the IP sequence of BFV and other foamy viruses (%) BFV-IP
, http://www.100md.com
8914-9413
9048-9413
9112-9413
9200-9413
BFV-FeFV
61.4
57.9
58.9
61.7
BFV-HFV
46.5
48.1
, 百拇医药
54.3
61.9
BFV-SFV1
44.1
46.4
52.0
58.1
BFV-SFV3
44.5
48.6
55.6
66.4
讨 论
, 百拇医药
本研究从本实验室分离并鉴定的BFV3026中国毒株中克隆到了完整的内部启动子,并构建了pBFVIP-luc表达质粒。体外瞬时表达实验表明,该启动子可以被牛泡沫病毒自身编码的反式激活因子ORF-1所激活,从而证实了牛泡沫病毒中内部启动子的存在。IP的存在使泡沫病毒的基因表达调节机制更显复杂。以前的一些研究表明:HFV的env IP区域为8 921~9 332,TATA盒位于转录起始点上游的9 164~9 169,SFV-1 env IP位于8 183~8 370,其Tas调控位点位于8 249~8 370,TATA盒为8 380~8 385。SFV3 env IP区域为9 534~9 820,9 731~9 736为其TATA盒。而本文克隆的BFV env 3′区段(基因组8 572~9 509)中包含了完整的牛泡沫病毒内部启动子。该启动子TATA盒位于基因组9 282~9 285之间。泡沫病毒的TATA盒序列及附近序列十分保守,均为TATAAAAGA*CA*AT。该TATA盒很可能与TATA盒结合因子TFIID相互作用,而该因子是宿主细胞RNA聚合酶Ⅱ复合物起始转录所必需的。核苷酸序列同源分析表明,BFV env基因3′端与HFV、SFV-1、SFV-3、FeFV同源性均达44%以上。其中与FeFV的同源性最高,达61.4%。但当区段逐渐向TATA盒靠近时,同源性逐步升高(表1)。这暗示着越靠近TATA盒序列保守性越强,很可能在TATA盒近上游包含有重要的调控位点或ORF-1反式激活的靶位点(TREIP)。但仅根据同源性分析还难以作出定论,而有待于对该区段进一步缺失或定点突变来确定TREIP的位置。
, 百拇医药
BFV基因组中存在两个启动子的意义有待于进一步阐明,与其它病毒相比,泡沫病毒中内部启动子的存在并非独特,鼠乳腺肿瘤病毒(mouse mammary tumor viruses)和人T细胞白血病病毒1型(human T-cell leukemia virus type 1)亦可以从LTR及env基因内部起始转录,但这些病毒的内部启动子不能被相应病毒编码的某种蛋白反式激活。而BFV IP及LTR均强列依赖牛泡沫病毒自身编码的反式激活因子ORF-1。尽管这两种启动子均能被ORF-1反式激活,但调控机制却不尽相同。将BFV IP TATA附近区域与LTR相应区段,以及BFV IP TATA盒上游区域与LTR U3区内的两个TRELTR进行比较(TREI:-983/-668,TREII:-470/-140,转录起始位点为+1)[8],均只发现极短簇的同源区。IP和LTR较低的同源性,暗示着两个启动子的反式激活可能有不同的因子参与。Mergia在对SFV-1的研究中曾提出过“时序模式(temporal pattern)”[5]。该模式认为,和许多DNA病毒一样,泡沫病毒在复制周期的不同时期,由不同的阶段特异性启动子(stage-specific promoter)控制不同的基因表达。即表达受2个或多个启动子调控。在病毒感染的早期,首先由IP起始转录,表达出病毒的调节蛋白Tas;随后,Tas一方面作用于LTR,表达出病毒结构蛋白,完成病毒颗粒的组装;另一方面反馈激活IP,合成更多的调节蛋白。由于至今还没有在泡沫病毒中发现与Rev/Rex相当的转录后调节蛋白,所以IP的存在在功能上替代了其它反转录病毒的转录后调节蛋白的作用。本文瞬时表达分析的结果也正好符合该模式,无BFV ORF-1存在时,即相当于病毒感染的早期,BFV IP的基础活性远远高于LTR,能够引发少量调节蛋白的表达;而有ORF-1存在时,即相当于病毒凭借IP的基础活性表达出部分调节蛋白后,IP的活性和LTR的活性均被大大激活。这一结果支持了“时序模式”,揭示了BFV与HFV、SFV-1、SFV-3有着相似的基因表达调控机制。
, 百拇医药
基金项目:国家自然科学基金(39770032);教育部博士点基金资助项目(98005516)
作者简介:张莉(1973-),女,湖南人,分子遗传学硕士。
联系作者:耿运琪
参考文献:
[1] Holzschu D L, Delaney M A, Renshaw R W, et al. The nucleotide sequence and spliced pol mRNA levels of the nonprimate spumavirus, bovine foamy virus [J] . J Virol, 1998, 72 (3) : 2177-2182.
[2] Lochelt M, Flugel R M, Aaboud M, et al. Human foamy virus genome possesses an internal, Bel-1-dependent and functional promoter [J] . Proc Natl Acad Sci USA, 1993, 90: 7317-7321.
, 百拇医药
[3] Lochelt M, Muranyi W, Flugel R M, et al. The human foamy virus internal promoter directs the expression of the functional bel-1 transactivator and bet protein early after infection [J] . J Virol, 1994, 68 (20) : 638-645.
[4] Mergia A. Simian foamy virus type 1 contains a second promoter located at the 3' end of the env gene [J] . Virology, 1994, 199: 219-222
[5] Campbell M, Renshaw G L, Renne R, et al. Characterization of the internal promoter of simian foamy viruses [J] . J Virol, 1994, 68 (8) : 4811-4820
[6] 刘淑红,陈荷新,陈家童,等. 牛泡沫病毒(BSV)3026毒株的分离及分子生物学鉴定[J].病毒学报,1997,13(2):140-145.
[7] 刘国文,纪永刚,梁臣,等.萤火虫荧光素酶基因构建BIV-LTR启动子表达体系[J].病毒学报,1994,10(4):377-380.
[8] 刘佳建,王世珍,张莉,等.牛泡沫病毒调节蛋白功能及其在LTR上应答元件的研究[J].病毒学报,2000,16(2):141~149.
来稿日期:1999-02-01;修回日期:1999-11-29, http://www.100md.com