œΏΝΘΧε‘ΎΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡΉς”Ο
Ής’ΏΘΚΚΈ–ßΕΪ
ΒΞΈΜΘΚΚΈ–ßΕΪΘΚΫβΖ≈ΨϋΒΎ“Μ“Ϋ‘ΚΗΈΒ®ΆβΩΤ÷––ΡΘ§άΦ÷ί730030 Ε≠Φ“Κη ≤ΧΨΑ–όΘΚΒΎ»ΐΨϋ“Ϋ¥σ―ßΈςΡœ“Ϋ‘ΚΗΈΒ®ΆβΩΤ÷––ΡΘ§÷Ί«λ 400038
ΙΊΦϋ¥ ΘΚΗΈ‘ύΦ≤≤ΓΘΜœΏΝΘΧεΘΜ»±―Σ-‘ΌΙύΉΔΥπ…Υ
œΏΝΘΧε‘ΎΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡΉς”ΟΚΈ–ßΕΪ Ήέ ω Ε≠Φ“Κη ≤ΧΨΑ–ό …σ–Θ
’Σ“ΣΘΚΗΈ‘ύΆβΩΤΓΔ–ίΩΥΒ»≤ΓάμΙΐ≥Χ÷–ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ≤ΜΩ…±ήΟβΘ§ΗΟΙΐ≥Χ“‘»±―ΣΓΔ»±―θΓΔΡήΝΩΙ©”Π÷–ΕœΈΣ ΦΕ·“ρΥΊΘ§Ψ≠œΗΑϊΗΤ≥§‘ΊΓΔ―θΉ‘”…Μυ–Έ≥…ΦΑΑΉœΗΑϊΫΰ»σ»ΐΗω÷Ί“ΣΜΖΫΎΘ§Ά®ΙΐœΗΑϊ“ρΉ”―Ή–‘Ϋι÷ ΒΡ ΆΖ≈Θ§≤ΜΒΪΕ‘»±―ΣΗΈ‘ύ‘λ≥…Υπ…ΥΘ§Εχ«“Ε‘»Ϊ…μΤδΥϋ‘ύΤς‘λ≥…“ΜΕ®ΒΡ…ΥΚΠΘΜ‘Ύ’β“Μ≤ΓάμΙΐ≥Χ÷–Θ§œΏΝΘΧεΕ‘œΗΑϊΡΎΗΤΒΡΒςΫΎΓΔ―θΉ‘”…Μυ–Έ≥…Β»ΨΏ”–Ζ«≥Θ÷Ί“ΣΒΡΉς”ΟΓΘ»γΚΈΖά÷ΈΗΈ‘ύ»±―Σ‘ΌΙύΉΔ ±œΏΝΘΧεΥπ…ΥΫΪ «ΫώΚσ Β―ι―–ΨΩΒΡ÷ΊΒψΓΘ
÷–ΆΦΖ÷άύΚ≈ΘΚR 575 ΈΡœΉ±ξ Ε¬κΘΚA ΈΡ’¬±ύΚ≈ΘΚ1007-8622(1999)04-0295-03
, http://www.100md.com
ΗΈ‘ύ «ΧεΡΎΉν¥σΒΡ¥ζ–ΜΤςΙΌΘ§œΏΝΘΧε «œΗΑϊ¥ζ–ΜΒΡ÷––Ρ≥ΓΥυΘ§œΏΝΘΧεΒΡ’ΐ≥ΘΚτΈϋΙΠΡήΚΆATP…ζ≥…Ε‘Ήι÷·ΤςΙΌΙΠΡήΦΑœΗΑϊΫαΙΙΒΡΆξ’ϊ–‘÷ΝΙΊ÷Ί“ΣΓΘΒΪœΏΝΘΧεΕ‘»±―ΣΓΔ»±―θΖ«≥ΘΟτΗ–Θ§»±―ΣΓΔ»±―θΒΡΗΈΉι÷·‘Ύ―ΣΝςΜ÷Η¥ΜρΗ¥―θΚσΤδΥπ…Υ≥ΧΕ»≤ΜΒΪΟΜ”–ΗΡ…ΤΘ§Ζ¥ΕχΫχ“Μ≤ΫΦ”÷ΊΘ§’β”κœΗΑϊΗΤ≥§‘ΊΓΔ―θΉ‘”…ΜυΒ»“ρΥΊ”–÷±Ϋ”ΒΡΙΊœΒ[1]ΓΘœΏΝΘΧε «ΝΣœΒΗΤ≥§‘ΊΓΔ―θΉ‘”…Μυ–Έ≥…ΚΆœΗΑϊΥάΆωΒΡ÷––ΡΜΖΫΎΓΘΫϋΡξΕ‘œΏΝΘΧε‘ΎΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡΉς”Ο―–ΨΩΫœΕύΘ§Ϋχ’ΙΫœΩλΘ§±ΨΈΡΨΆ’βΖΫΟφΒΡΡΎ»ίΉω“ΜΉέ ωΓΘ
1 œΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ”κ»±―Σ‘ΌΙύΉΔΗΤ≥§‘ΊΒΡΝΣœΒ
ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΩ…÷¬œΗΑϊΗΤ≥§‘ΊΘ§ «“ΐΤπΗΈœΗΑϊΥπ…ΥΒΡ‘≠“ρ÷°“ΜΘ§ΕχœΏΝΘΧεΨΏ”–«Ω¥σΒΡΨέΜΐCa2+ΡήΝΠΘ§Ε‘ΗΤάκΉ”Έ»Χ§ΤπΒΫ“ΜΕ®ΒΡΒςΫΎΉς”ΟΘ§ΒΪœΏΝΘΧε…ψCa2+ΡήΝΠ”–œόΘ§»τΥπ…Υ“ρΥΊ≤ΜΡή«ε≥ΐΘ§Ψ≠Ης÷÷ΆΨΨΕΉν÷’Ω…÷¬œΗΑϊΡΎΗΤ≥§‘Ί[2Θ§3]ΓΘ
ΫϋΡξΕ‘œΗΑϊΜΒΥάΒΡ―–ΨΩ±μΟςΘ§œΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ(Mitochondrial membrane permeability transition,MMPT)‘ΎœΗΑϊΥάΆω÷–ΤπΉ≈ΙΊΦϋΒΡΉς”Ο[4]ΓΘ‘ΎΗς÷÷Υπ…Υ“ρΥΊΒΡΉς”Οœ¬Θ§MMPT“ΜΒ©ΖΔ…ζΘ§œΏΝΘΧεΡΛΆ®ΆΗ–‘―ΗΥΌ…ΐΗΏΘ§œΗΑϊΥάΆωΥφ÷°ΖΔ…ζΓΘ“÷÷ΤMMPTΖΔ…ζΩ…―”ΜΚœΗΑϊΥάΆω[5]ΓΘΡΩ«Α»œΈΣMMPTΒΡΖΔ…ζΜζ÷ΤΑϋά®œΏΝΘΧεΡΎΡΛΒΡΖ«ΧΊ“λ–‘Υπ…ΥΚΆ“Μ÷÷ΨόΆ®Βά(Megapore)ΒΡΩΣΖ≈[4Θ§6]ΓΘΕχœΏΝΘΧεΡΎΡΛΖ«ΧΊ“λ–‘Υπ…ΥΩ…”…ΗΤ≥§‘Ί¥ΞΖΔΒΡ―θΉ‘”…ΜυΥπ…ΥΒ»‘λ≥…ΓΘΙΊ”ΎΒΦ÷¬MMPTΒΡΧΊ βΜζ÷ΤΒΡ―–ΨΩ±μΟςΘΚœΏΝΘΧεΜυ≤ψ÷–Ca2ΓFΒΡ¥σΝΩΨέΜΐ’ΐ «ΨόΆ®Βά(Megapore)ΩΣΖ≈ΒΡ‘≠“ρ÷°“Μ[5Θ§6]ΓΘΙ œΏΝΘΧεΗΤ≥§‘ΊΆ®ΙΐMMPTΩ…ΒΦ÷¬œΗΑϊ÷¬Υά–‘Υπ…ΥΓΘ
, http://www.100md.com
Ε‘ΨόΆ®ΒάΫχ“Μ≤Ϋ―–ΨΩΖΔœ÷Θ§œΏΝΘΧεΡΛ…œΒΡΨόΆ®ΒάΨΆ «÷±ΨΕΈΣ20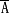 (2 nm)ΒΡCa2+“άάΒ–ΓΩΉΘ§ΗΟ–ΓΩΉΨΏ”–―Γ‘ώ–‘Ά®ΆΗΉς”Ο[7]ΓΘ“―ΦχΕ®≥ωΗΟ–ΓΩΉΈΣœΏΝΘΧεΡΎΡΛΒΡΗΏΒγΒΦ–‘ΓΔœύΒ±ΒΆάκΉ”ΧΊ“λ–‘¥σΆ®ΒάΓΘœΏΝΘΧεΡΎCa2+ΚΆPiΒΡ…ΐΗΏΜρΜν–‘―θ¥ΧΦΛ ±–ΓΩΉΦ¥ΩΣΖ≈[6]Θ§Β±œΏΝΘΧεCa2+±Μ»Ξ≥ΐΚσ–ΓΩΉ―ΗΥΌΙΊ±’[4]ΘΜ…ζάμΦΝΝΩΒΡATP(>1mM)Ρή“÷÷ΤΤδΩΣΖ≈ΓΘ’ΐ≥Θ«ιΩωœ¬Θ§–ΓΩΉ≥…ΖίΜΖ«ΉΚΆΥΊ(cyclophilin)Έ§≥÷ΤδΩΣΖ≈ΓΘΜΖφΏΥΊA(cyclosporin A,CSA)Ρή”κΜΖ«ΉΚœΥΊΫαΚœ Ι–ΓΩΉΙΊ±’Θ§Ά§ ±Θ§CSAΨΏ”––ό≤ΙœΏΝΘΧεΡΎΡΛΒΡΙΠΡή(patch-clamp technique)[5,7].¥ΥΆβΘ§–ΓΩΉΒΡΩΣΖ≈ΜΙ”κœΏΝΘΧεΜυ÷ ΒΡpH”–ΙΊΘ§Β±œΏΝΘΡΎpH≥ Υα–‘ ±–ΓΩΉΒΡΩΣΖ≈Ιΐ≥Χ±δ¬ΐΓΘΝμΆβΘ§œΏΝΘΧεΆ®ΆΗ–‘ΗΡ±δΩ…”…œΏΝΘΧεΡΛΝΉ÷§ΒΡ»ΞθΘΜ·-‘ΌθΘΜ·―≠ΜΖΈ…¬““ΐΤπΓΘ
(2 nm)ΒΡCa2+“άάΒ–ΓΩΉΘ§ΗΟ–ΓΩΉΨΏ”–―Γ‘ώ–‘Ά®ΆΗΉς”Ο[7]ΓΘ“―ΦχΕ®≥ωΗΟ–ΓΩΉΈΣœΏΝΘΧεΡΎΡΛΒΡΗΏΒγΒΦ–‘ΓΔœύΒ±ΒΆάκΉ”ΧΊ“λ–‘¥σΆ®ΒάΓΘœΏΝΘΧεΡΎCa2+ΚΆPiΒΡ…ΐΗΏΜρΜν–‘―θ¥ΧΦΛ ±–ΓΩΉΦ¥ΩΣΖ≈[6]Θ§Β±œΏΝΘΧεCa2+±Μ»Ξ≥ΐΚσ–ΓΩΉ―ΗΥΌΙΊ±’[4]ΘΜ…ζάμΦΝΝΩΒΡATP(>1mM)Ρή“÷÷ΤΤδΩΣΖ≈ΓΘ’ΐ≥Θ«ιΩωœ¬Θ§–ΓΩΉ≥…ΖίΜΖ«ΉΚΆΥΊ(cyclophilin)Έ§≥÷ΤδΩΣΖ≈ΓΘΜΖφΏΥΊA(cyclosporin A,CSA)Ρή”κΜΖ«ΉΚœΥΊΫαΚœ Ι–ΓΩΉΙΊ±’Θ§Ά§ ±Θ§CSAΨΏ”––ό≤ΙœΏΝΘΧεΡΎΡΛΒΡΙΠΡή(patch-clamp technique)[5,7].¥ΥΆβΘ§–ΓΩΉΒΡΩΣΖ≈ΜΙ”κœΏΝΘΧεΜυ÷ ΒΡpH”–ΙΊΘ§Β±œΏΝΘΡΎpH≥ Υα–‘ ±–ΓΩΉΒΡΩΣΖ≈Ιΐ≥Χ±δ¬ΐΓΘΝμΆβΘ§œΏΝΘΧεΆ®ΆΗ–‘ΗΡ±δΩ…”…œΏΝΘΧεΡΛΝΉ÷§ΒΡ»ΞθΘΜ·-‘ΌθΘΜ·―≠ΜΖΈ…¬““ΐΤπΓΘ
, http://www.100md.com
ΒΪΝμ”–―–ΨΩ»œΈΣΘ§‘ΎœΗΑϊΥπ…Υ÷–Θ§œΏΝΘΧε…ψΗΤ‘ωΕύ“≤–μ «ΦΧΖΔ–‘ΗΡ±δΒΡΫαΙϊΘ§ΕχœΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ(MMPT)≤≈ « ΦΕ·“ρΥΊΘ§“ρΕχΕ‘ΑϊΫ§ΗΤ≥§‘Ί‘ΎΉι÷·»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡ÷Ί“Σ–‘Χα≥ω÷ “…Θ§[8]Θ§“ρΈΣ‘Ύ¥σ σΗΈ‘ύ»±―θ35 min‘ΌΙύΉΔ10 minΒΡΡΘ–Ά÷–Θ§ΗΈΉι÷·ΒγΨΒœ¬Ά§“Μ ”“ΑΒΡΝΫΗωΗΈœΗΑϊ÷–Θ§“ΜΗωœΏΝΘΧε’ΐ≥ΘΘ§ΕχΝμ“ΜΗω‘ρΥ°÷Ή±δ–‘Θ§Κσ’Ώ“≤–μ“―”–œΏΝΘΧεΆ®ΆΗ–‘ΗΡ±δΘΜΕχ÷ΒΒΟΉΔ“βΒΡ «¥Υ ±»‘Έ¥ΦϊΤδΥϋœΗΑϊΤς”––ΈΧ§―ßΖΫΟφΒΡ±δΜ·[11]ΓΘ’βΫω «≥θ≤ΫΙέ≤λΒΡΫαΙϊΘ§“Σ≤ϊΟςΤδœξœΗΜζ÷ΤΘ§ΜΙ”–¥ΐΗϋΫχ“Μ≤Ϋ―–ΨΩΓΘ
2 œΏΝΘΧε‘Ύ―θΉ‘”…Μυ–Έ≥…÷–ΒΡΉς”Ο
ΜΤύ―Ώ ―θΜ·ΟΗΚΆ÷––‘ΑΉœΗΑϊ «―θΉ‘”…ΜυΒΡΝΫΗω÷Ί“Σά¥‘¥Θ§[9]Θ§»±―ΣΚΆ‘ΌΙύΉΔΨυΩ…≤ζ…ζΫœΕύ≥§―θΜ·ΈοΘ§“ΐΤπΉι÷·Υπ…ΥΓΘœΏΝΘΧε «ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΤΎΦδΜν–‘―θ≤ζ…ζΒΡ÷Ί“ΣΤςΙΌ[10]ΓΘ
2.1 ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΜν–‘―θ ΆΖ≈ΧΊΒψ”κΗΈœΗΑϊΥπ…ΥΒΡΙΊœΒΘΚΕ‘ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ―–ΨΩ±μΟςΘ§Η¥―θΚσΜν–‘―θ ΆΖ≈ ±œύ”κΗΈœΗΑϊΥπ…ΥΟή«–œύΙΊΓΘJaeschkeΒ»[11]ΉηΕœ≤ΩΖ÷ΗΈ‘ύ―ΣΝς45 minΚσ‘ΌΙύΉΔ(RP)≤ΜΆ§ ±ΦδΘ§ΖΔœ÷ΗΈΉι÷·≥ωœ÷ΝΥΥΪœύΥπ…ΥΓΘ‘ΎΜ÷Η¥ΙύΉΔΒΡ1 hΡΎΘ§―ΣΫ§ΉΣΑ±ΟΗΜν–‘…ΐΗΏΘ§¥ΥΈΣΗΈ‘ύ»±―Σ‘ΌΙύΉΔΒΡΔώœύΥπ…Υ(initial injury)ΘΜΫ”œ¬»ΞΒΡ5 h Θ§ΉΣΑ±ΟΗΒΡΜν–‘Έ§≥÷‘ΎΔώœύΥπ…ΥΥ°ΤΫΕχ‘ΎRPΚσ6ΓΪ24 hΘ§ΉΣΑ±ΟΗΜν–‘‘Ό¥Έœ‘÷χ…ΐΗΏΘ§±μΟς»±―Σ‘ΌΙύΉΔΗΈ‘ύΜΙ¥φ‘ΎΔρœύΥπ…Υ(progressive injury)ΓΘ¥”ΕχΩ…“‘Ω¥≥ωΗ¥―θ ±ΗΈœΗΑϊΥπ…Υ”κΜν–‘―θΒΡ ΆΖ≈ ±œύΨΏ”–÷±Ϋ”ΝΣœΒΓΘ Β―ιΜΙ÷ΛΟςRPΗΈ‘ύΒΡΔώœύΥπ…Υ”κΗΈώΦΩίΖώ œœΗΑϊ(KC)”–ΙΊΓΘKCΩ…“ρΗΤ≥§‘ΊΓΔΆΧ …Ής”ΟΜρ≤ΙΧεœΒΆ≥ΕχΦΛΜνΘ§ΜνΜ·ΒΡKC≥ΐ÷ς“Σ ΆΖ≈Μν–‘―θΆβΘ§ΜΙΩ… ΆΖ≈ΒΑΑΉΟΗΦΑΗς÷÷œΗΑϊ“ρΉ”Θ§ΫιΒΦΕ‘ΗΈœΗΑϊΒΡΕΨ–‘Ής”ΟΘ§ΒΦ÷¬―œ÷ΊΒΡΗΈ‘ύ‘ΌΙύΉΔΥπ…ΥΘΜΗ…»≈KCΙΠΡήΒΡ“©ΈοΨΏ”–Οςœ‘ΒΡ÷ΈΝΤΉς”Ο[12Θ§13]Θ§ΕχΔρœύΥπ…Υ”κΑΉœΗΑϊ‘ΎΨ÷≤ΩΒΡΨέΦ·ΒΫ“ΜΕ®ΝΩ≤ΔΦΛΜνΚσ ΆΖ≈ΕΨ–‘Ϋι÷ ”–ΙΊΘ§ΨΓΙήΑΉœΗΑϊ‘ΎRP≥θΤΎ‘ΎΗΈώΦ”κΡΎΤΛœΗΑϊ’≥ΗΫ‘ω«Ω≤ΔΩΣ ΦΨέΦ·Θ§ΒΪ≤Δ≤Μ≤Έ”κΗΈΉι÷·ΔώœύΥπ…Υ[13]ΓΘΆ§ ±ΜΙΖΔœ÷KCΒΡΔώœύΥπ…ΥΩ…ΦΛΜν÷––‘ΑΉœΗΑϊ(PMN)Θ§ ΙΤδ ΆΖ≈Μν–‘―θΦΑΒΑΑΉΟΗΒ»ΕΨ–‘Ϋι÷ ΙΞΜςΗΈœΗΑϊΘ§ΜΒΥάΗΈœΗΑϊΖ¥Ιΐά¥”÷Ω…¥ΌΫχ≤ΙΧεΦΛΜ·ΚΆPMNΜνΜ·Θ§»γ¥ΥΕώ–‘―≠ΜΖΘ§Ήν÷’ΒΦ÷¬―œ÷ΊΗΈœΗΑϊΥπ…ΥΓΘΒΞΩΥ¬ΓΩΙΧεΩ…Φθ…ΌPMN’≥ΗΫ≤Δ Ι‘ΌΙύΉΔΥπ…ΥΦθ«αΓΘ[14]ΓΘ
, http://www.100md.com
2.2 Μν–‘―θ”κ»±―Σ‘ΌΙύΉΔΚσΥπ…ΥΗΈœΗΑϊ‘ΎΗΈœΌ≈ί÷–ΒΡΖ÷≤ΦΘΚ“‘«Α―–ΨΩΖΔœ÷Θ§ΗΈ“Ε«–≥ΐΚσΗΈœΗΑϊΒΡ‘Ό…ζ÷ς“ΣΖΔ…ζ‘ΎΗΈ≈ίΔώ¥χΘ§ΕχΫα‘ζΕ·ΈοΗΈ―ΣΙή ΙΗΈ‘ύ»±―θΜρ”Σ―χ≤ΜΝΦ–‘‘γΤΎΗΈ”≤Μ·ΥυΖΔ…ζΒΡΗΈΥπΚΠ÷ς“ΣΖΔ…ζ‘ΎΔσ¥χΘ§’βΩ…Ϋβ ΆΈΣΔώ¥χΗΈœΗΑϊΩΩΫϋ÷–÷α―ΣΙήΘ§”≈œ»ΜώΒΟΗΜ”Ύ”Σ―χΒΡΈο÷ ΚΆΚ§―θ≥δΉψΒΡ―Σ“ΚΘ§“ρ¥Υ”≈œ»‘Ό…ζΘ§ΕχΔσ¥χΉι÷·PO2ΉνΒΆΘ§Ι Ήν»ί“Ή≥ωœ÷»±―θ–‘ΗΈΥπΚΠΚΆΗΈœΗΑϊΥάΆωΓΘ
Ρ«Ο¥Θ§ΗΈ‘ύ»±―ΣΚσ‘ΌΙύΉΔΥπ…ΥΒΡΧΊΒψ « ≤Ο¥ΡΊ?FlreemanΒ»ΒΡ―–ΨΩ»œΈΣΘΚΜΙ‘≠–ΆΙ»κΉΗ κΡ(GSH)‘ΎΗΈ‘ύ «÷Ί“ΣΒΡΉ‘”…Μυ«ε≥ΐΦΝΓΘ―–ΨΩΖΔœ÷Θ§¥σ σΗΈ‘ύ»±―Σ90 minΚσ‘ΌΙύΉΔΘ§ΗΈΉι÷·GSHΒΆ’Ώ(»γΫϊ ≥)ΗΈ‘ύΥπΚΠ≥ΧΕ»ΫœGSHΗΏ’ΏΗϋΈΣ―œ÷ΊΘ§≤Δ«“ΥπΚΠΗΈœΗΑϊ÷ς“ΣΖ÷≤Φ‘Ύ–Γ“ΕΦδΟ≈Ψ≤¬ω÷ήΈß«χ”ρ(Φ¥Δώ¥χ)Θ§’β÷÷±μœ÷‘ΎGSHΆξ»ΪœϊΚΡ’ΏΗϋΦ”Οςœ‘Θ§“ρΕχ÷ß≥÷―θΉ‘”…Μυ“ΐΤπΗΈΥπ…Υ’β“Μ¬έΕœ[15]ΓΘΆ§ ± Β―ι“≤÷ΛΟςΘ§”Ο¬»Μ·νή¥ΠάμΫϊ ≥ΒΡΕ·ΈοΘ§“‘‘ωΦ”GSH‘ΎΗΈΉι÷·ΒΡΚ§ΝΩΘ§ΫαΙϊ»±―ΣΚσΗΈœΗΑϊΜΒΥά≥ΧΕ»Οςœ‘Φθ«α[15]ΘΜ’β÷÷Ε‘»±―Σ‘ΌΙύΉΔΥπ…ΥΒΡ«χ¥χ–‘≤ν“λΘ§Ω…Ρή”κ“‘œ¬“ρΥΊ”–ΙΊΘΚΔΌ”κKupfferœΗΑϊ‘ΎΗΈώΦΒΡ≤ΜΨυ÷ –‘Ζ÷≤Φ”–ΙΊΓΘ¥σ σΗΈKupfferœΗΑϊ‘Φ’ΦΗΈ Β÷ ΒΡ2.1%Θ§ΒΪ“‘ΗΈ–Γ“Ε÷ή±Ώ¥χΖ÷≤ΦΫœΕύΓΘkupfferœΗΑϊΒΡΜνΜ·‘ΎΜζΧεΖάΈάΜζΡή÷–”–ΜΐΦΪΒΡΉς”ΟΘ§ΒΪΜνΜ·ΚσΒΡKupfferœΗΑϊ“‘ΤδΖ÷ΟΎΒΡΗς÷÷œΗΑϊ“ρΉ”(cytotines)Ής”Ο”ΎΗΈœΗΑϊΓΔΝΎΫϋΉι÷·ΦΑ»Ϊ…μΓΘ»±―ΣΓΔ‘ΌΙύΉΔΩ…÷±Ϋ”ΜρΦδΫ”ΦΛΜνKupfferœΗΑϊ(Μρ÷––‘ΑΉœΗΑϊ)Θ§Ω… ΆΖ≈Εύ÷÷œΗΑϊ“ρΉ”ΚΆΫι÷ [16]Θ§Τδ÷–Αϋά®O2¥ζ–Μ÷–Φδ≤ζΈοH2OΓΔO?2ΓΔOH?Ή‘”…ΜυΓΔO2ΦΑTNFΒ»ΓΘΔΎ”κ»±―ΣΚσΓΑΈόΙύΉΔΓ±(no-reflow)ΜρΓΑΈό―Σ‘ΌΙύΉΔΓ±(reflow-paradox)“ΐΤπΈΔ―≠ΜΖΈ…¬“”–ΙΊ [17]ΓΘΗΈώΦ «ΧΊΜ·ΒΡΟΪœΗ―ΣΙήΘ§Τδ÷±ΨΕ”κΤδΥϋΟΪœΗ―ΣΙήœύΥΤΘ§ΒΪΟΩΗωΗΈœΗΑϊΫτΝΎΝΫ…œΗΈώΦΘ§“ρΕχΕ‘ΟΪœΗ―ΣΙήΉη»ϊΒΡΡΆ ήΫœ«ΩΘ§‘Ύ…ζάμ«ιΩωœ¬“≤≥ΘΖΔ…ζΒΞΗω―ΣώΦΒΡΉη»ϊΓΘΒΪΗΈ‘ύ≥Λ ±Φδ»±―ΣΚσ”…”ΎΡΎΤΛΥ°÷ΉΓΔ―ΣώΦΡΎ―Σ“Κ≈®ΥθΦΑΈΔΨ≤¬ωΉη»ϊΒ»Θ§ Ι‘ΌΙύΉΔΚσ»±―Σ»±―θ»‘‘ΎΦΧ–χ(no-reflow)Θ§ΒΦ÷¬‘ΌΙύΉΔ≤Μ‘»Θ§Εχ‘ΌΙύΉΔΚσΓΑΈό―Σ‘ΌΙύΉΔΓ±(reflow-paradox) «”…”ΎΗ¥ΝςΜρΗ¥―θΦΛΜνΝΥΑΉœΗΑϊΘ§Ψ≠œΗΑϊ±μΟφ’≥ΗΫΖ÷Ή”ICAM-1ΓΔCD11/CD18ΓΔLΓΔP-―Γ‘ώΥΊ”κΡΎΤΛœΗΑϊάΈΙΧ’≥ΗΫΕχ“ΐΤπΗΈώΦΉη»ϊΓΘΆ®Ιΐ“‘…œΝΫ÷÷ΖΫ ΫΘ§ ΙΗ¥ΝςΚσΜν–‘―θ ΆΖ≈‘ωΕύ≤Δœό÷Τ‘ΎΨ÷≤ΩΘ§Ά§ ±Θ§”…”Ύ―θΉ‘”…ΜυΤΫΨυ ΌΟϋΚήΕΧΘ§Ϋω10?3Οκ[18]Θ§‘ΎΈΔ―≠ΜΖΈ…¬“ΒΡ«ιΩωœ¬≤ΜΡήΝΔΦ¥‘ΎΗΈΉι÷·≤Ξ…ΔΜρ¥σΝΩ ΆΖ≈»κ―Σ―≠÷–Θ§‘λ≥…Ψ÷≤ΩΉι÷·―œ÷ΊΥπ…Υ(»γΟ≈Ιή«χ)Θ§Rymsa―–ΨΩΖΔœ÷Θ§RP≥θΤΎ‘ΎΟ≈¬ω÷ήΈß«χ”ρΖ÷άκ≥ωΨΏ”–Κή«ΩΆΧ …Μν–‘ΒΡ¥σKCΘ§ΤδΉ‘ΖΔ≤ζ…ζΒΡO?2‘ωΦ”3±Ε“‘…œΓΘSuzukiΒ»‘ΎΗΈ‘ύ»±―Σ90 min‘ΌΙύ60 minΒΡΉι÷·―ßΙέ≤λ÷–ΖΔœ÷Θ§≤Γ±δ÷ς“ΣΖ÷≤Φ‘ΎΗΈ≈ίΔώ¥χ÷ΝΔρ¥χΘ§Ω…ΦϊΗΈœΗΑϊ―œ÷Ί±δ–‘Ή«÷ΉΓΔΗΈώΦπω―Σ[9]ΓΘΙΊ”ΎΤδΥπ…ΥΖ÷≤Φ≤ΩΈΜΒΡ»Ζ«–Μζ÷ΤΜΙ”–¥ΐΫχ“Μ≤Ϋ―–ΨΩΓΘ
, ΑΌΡ¥“Ϋ“©
ΒΪGonzalez-Flecha[15]―–ΨΩΖΔœ÷Θ§ΗΈœΌ≈ίΔρΓΔΔσ¥χΕ‘»±―Σ‘ΌΙύΉΔΥπ…Υ±»ΫœΟτΗ–Θ§ΡΩ«Α―–ΨΩ»œΈΣΗΈ≈ίΈό¬έ‘ΎœΗΑϊΤςΫαΙΙΜρΟΗΒΡΉιΖίΕΦ”–Υυ≤ΜΆ§Θ§’β «ΥϋΕ‘»±―Σ‘ΌΙύΉΔΥπ…ΥΟτΗ––‘≤ΜΆ§ΒΡΜυ¥ΓΓΘΡΩ«ΑΕ‘ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…ΥΟτΗ––‘ΒΡΉι÷·―ßΕ®ΈΜ»‘”–’υ“ιΘ§ΜΙ”–¥ΐΫχ“Μ≤ΫΧΫΧ÷ΓΘ
3 »±―Σ‘ΌΙύΉΔœΏΝΘΧεΚτΈϋΜν–‘ΒΡ±δΜ·
«ΑΟφ“―Ψ≠ΧαΒΫœΏΝΘΧε‘ΎœΗΑϊΗΤ≥§‘ΊΦΑ―θΉ‘”…Μυ–Έ≥…÷–ΒΡΒςΫΎΉς”ΟΓΘΒΪΒ±’β÷÷ΒςΫΎΉς”Ο≥§Ιΐ“ΜΕ®œόΕ»Θ§±Ί»Μ“ΐΤπœΏΝΘΧεΉ‘…μΥπΚΠΘ§’β÷÷ΥπΚΠ≥ωœ÷«Α Ήœ»≥ωœ÷“ΜœΒΝ–¥ζ≥Ξ–‘ΗΡ±δΘ§Ι ΝΥΫβΤδ±δΜ·Ιφ¬…Ε‘»±―Σ‘ΌΙύΉΔΥπ…ΥΒΡΖά÷ΈΨΏ”–ΜΐΦΪΒΡΉς”ΟΓΘ
ΤάΦέœΏΝΘΧεΚτΈϋΜν–‘≥Θ”ΟΚτΈϋΩΊ÷Τ¬ (RCR)ΚΆΝΉ―θ±»(P/O)ά¥±μ ΨΓΘΥϋΟ« «Ζ¥”≥œΏΝΘΧεΆξ’ϊ–‘ΓΔ―θΜ·ΝΉΥαΜ·≈ΦΝΣ≥ΧΕ»ΦΑ―θΜ·ΝΉΥαΜ·–߬ ΒΡΝιΟτ÷Η±ξΓΘΗΈ‘ύ»±―Σ‘ΌΙύΉΔ ±œΏΝΘΧεΆ®ΆΗ–‘ΒΡΗΡ±δΓΔœΏΝΘΧεΡΎΟΗΚΆΗ®ΟΗΒΡΕΣ ßΘ§“‘ΦΑ”…¥Υ‘λ≥…ΒΡ―θΜ·ΝΉΥαΜ·’œΑ≠Θ§ΨυΩ…”ΑœλœΏΝΘΧεΚτΈϋΜν–‘ΓΘIsozaki‘Ύ―–ΨΩΦδΕœΦΑ≥÷–χΗΈ‘ύ»±―ΣΕ‘œΏΝΘΧεΙΠΡήΒΡ”ΑœλΚσΖΔœ÷Θ§ΗΈ‘ύ‘Ύ≥÷–χ»±―Σ30 minΚσΕ‘RCRΦΑP/O”Αœλ≤Μ¥σΘΜ»τΫΪ»±―ΣΉή ±Φδ…ηΕ®‘Ύ60 minΘ§Ψ≠Ιέ≤λΖΔœ÷ΦδΕœ»±―Σ’ΏœΏΝΘΧεΙΠΡήΟςœ‘ΚΟ”Ύ≥÷–χ»±―Σ’ΏΘ§±μΟςΥφ≥÷–χ»±―Σ ±ΦδΒΡ―”≥ΛΘ§œΏΝΘΧεΒΡΆξ’ϊ–‘ ήΒΫ“ΜΕ®ΒΡ”ΑœλΓΘ
, ΑΌΡ¥“Ϋ“©
KorokawaΒ»―–ΨΩΗΈ¥σ≤ΩΖ÷«–≥ΐΚσ≤–ΗΈœΗΑϊœΏΝΘΧεΒΡΙΠΡή ±ΦϊΒΫΘ§ ÷ θΚσΒΎ“ΜΧλCRCΦΑR3Ϋœ ÷ θ«ΑΟςœ‘‘ωΗΏΘ§2ΓΪ5 dΚσ÷πΫΞΫΒΒΆ÷Ν θ«ΑΥ°ΤΫΘ§ΥΒΟς≤–ΗΈ¥ζ–ΜΜν‘ΨœΏΝΘΧεΙΠΡή¥ζ≥Ξ–‘‘ω«ΩΓΘΒΪKobayashi[19]Ε‘»±―Σ‘ΌΙύΉΔ ±ΗΈ‘ύœΏΝΘΧεΚτΈϋΜν–‘Ιέ≤λΖΔœ÷Θ§»±―Σ120 minœΏΝΘΧεRCRΓΔR3Ϋœ»±―Σ«ΑΟςœ‘ΫΒΒΆΘ§‘ΌΙύΉΔΚσΥδ”–ΥυΧαΗΏΘ§ΒΪ»‘Οςœ‘ΒΆ”Ύ θ«ΑΥ°ΤΫΓΘGonzalez-Flecha[10]ΒΡ―–ΨΩ”κ“‘…œœύΥΤΘ§Ιέ≤λΝΥ»±―Σ60 minΓΔ120 minΦΑ180 minΦΑ‘ΌΙύΉΔ30 min ±œΏΝΘΧεΚτΈϋΙΠΡήΒΡΗΡ±δΘ§Φϊ»±―ΣΚσRCRΓΔR3Ψυ≤ΜΆ§≥ΧΕ»ΫΒΒΆΘ§≤ΔΥφ»±―Σ ±Φδ―”≥ΛΘ§’β÷÷±δΜ·‘ΫΟςœ‘Θ§R4‘ρΈόΟςœ‘±δΜ·ΘΜ‘ΌΙύΉΔ30 min ±Θ§“‘…œ÷Η±ξ≤ΜΒΪΟΜ”–…ΐΗΏΖ¥ΕχΫœ»±―ΣΚσΟςœ‘ΫΒΒΆΘ§ΒΪ»±―Σ180 min‘ΌΙύΉΔ30 minΉι‘ρ±δΜ·≤ΜΟςœ‘ΜρΈό±δΜ·Θ§“≤–μ¥Υ ±œΏΝΘΧε¥ζ≥Ξ“―ΒΫΦΪœόΘ§Ε‘Ης÷÷¥ΧΦΛΟτΗ––‘ΫΒΒΆΥυ÷¬ΓΘ
Ής’ΏΦρΫιΘΚΚΈ–ßΕΪ(1962-)Θ§Ρ–Θ§ΥΕ ΩΘ§÷ς÷Έ“Ϋ Π
, ΑΌΡ¥“Ϋ“©
≤ΈΩΦΈΡœΉΘΚ
[1]liu W,Schob O, Pugmire JE,et al.Glycohydrolases as markers of hepatic ischemia-reperfusion injury and recovery [J].Hepatology,1996,24:157.
[2]Hatanaka N,Kamijke W,Shimizu S,et al.Ca2ΓFrelease from mitochondrial induces cytosiolic enzyme leakage in anoxis liver [J].J Surg Res,1995,58:485.
[3]Masini A,Gallesi D,Giovannini F,et al. Membrane potential of hepatic mitochondrial after cute cocaine administration in rat-the role of mitochondrial reduced glutathione [J].Hepatology,1997,25:305.
, http://www.100md.com
[4]Saris N-E L,Eriksson KD.Mitochondrial dysfunction in ischemia-reperfusion [J].Acta Anaesthesiologica Scandinavia,1995,29:(suppl 107):171.
[5]Geexaerts MD,Ronveavx-Dupal MF,Lemasters JJ,et al.Cytosilic free Ca2+and proteolysis in lethal oxidative injury in endothelial cell[J].Am J Physiol,1991,261(5):C889.
[6]Szabo I, Zoratti M.The giant channel of the inner mitochondrial membrane is inhibited by cyclosporin A [J].J Biol,Chem,1991,266:3376.
, ΑΌΡ¥“Ϋ“©
[7]Bernard P,Vassanell S,Veronese P,et al.Modulation of the mitochondrial permeability transition pore[J].J Biol Chem,1992,267:2934.
[8]Jacobs WR,Sgambati M,Gomez G,et al.Role of cytosolic Ca 2+ in renal tubule damage induced by anoxia [J].Am J Physiol,1991,260:C545.
[9]Suzuki S,Toledo-pereyra LH,Rodriguez FL,et al. Neutrophil infilration as an important factor in liver ischemia and reperfusion injury[J].Transplantation,1993,55(6):1265.
, ΑΌΡ¥“Ϋ“©
[10]Gonzalez-Flecha B, Cutrin JC,Boveris A.Time course and mechanism of oxidative stress and tissue damage in rat liver subjected to in vivo ischemia-reperfusion[J]. J Clin Invest 1993,91:456.
[11]Jaeschke H,Farbood A,Smith W.Neutrophils contribute to ischemia/reperfusion injury in rat liver in vivo [J].FASEB J ,1990,4:3355.
[12]Brass CA,Roberts TG.Hepatic free radical production after cold storage;kupffer cell-dependent and independent mechanisms in rats [J].Gastroenterology,1995,108:1167.
, http://www.100md.com
[13]Niwano M,Arii S,Monden K,et al. Aneliovation of sinusoided endothelial cell damage by kupffer cell blockade during cell preservation of rat liver [J].J Surg Res,1997,72:36.
[14]Marubayashi S,Oshire Y,Maeda T,et al.Protective effect of monoclonal antibodies to adhesion molecules on rat liver ischemia-reperfusion injury [J].Surgery,1997,122:45.
[15]Jennische E. Possible influence of glutathine on postischemic liver injury [J].Acta Pathol Microbiol Scand,[C]1984,92:55.
, ΑΌΡ¥“Ϋ“©
[16]Shibuyo H,Ohkohchi N,Seya K,et al.Kupffer cell generate superoxide anions and modulate reperfusion injury in rat liver after cold preservation [J].Hepatology,1997,25:356.
[17]Menger MD.Microcirculatory disturbance secondary to ischemia-reperfusion [J].Transplantation,1995,27(5):2863.
[18]¬§γ≤ΤΦΘ§ Ζ≥¬»ΟΘ§Άτ ΚλΘ§Β».¥σ σΗΈ»±―Σ‘ΌΙύΝςΤΎΦδ÷§÷ Ιΐ―θΜ·Ή‘”…ΜυΕ·Χ§±δΜ·ΦΑ≤ΓάμΗΡ±δΆ§≤ΫΙέ≤λ.÷–ΜΣ Β―ιΆβΩΤ‘”÷ΨΘ§1992Θ§9ΘΚ11.
[19]Kobayashi H,Nonami T,Kurokawa T,et al.Mechanism and prevention of ischemia-reperfusion liver injury in rats [J] J Surg Res,1991,51:240.
, http://www.100md.com
’Ηε»’ΤΎΘΚ1999-01-20
œΏΝΘΧε‘ΎΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡΉς”Ο
ΚΈ–ßΕΪ Ήέ ω Ε≠Φ“Κη ≤ΧΨΑ–ό …σ–Θ
’Σ“ΣΘΚΗΈ‘ύΆβΩΤΓΔ–ίΩΥΒ»≤ΓάμΙΐ≥Χ÷–ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ≤ΜΩ…±ήΟβΘ§ΗΟΙΐ≥Χ“‘»±―ΣΓΔ»±―θΓΔΡήΝΩΙ©”Π÷–ΕœΈΣ ΦΕ·“ρΥΊΘ§Ψ≠œΗΑϊΗΤ≥§‘ΊΓΔ―θΉ‘”…Μυ–Έ≥…ΦΑΑΉœΗΑϊΫΰ»σ»ΐΗω÷Ί“ΣΜΖΫΎΘ§Ά®ΙΐœΗΑϊ“ρΉ”―Ή–‘Ϋι÷ ΒΡ ΆΖ≈Θ§≤ΜΒΪΕ‘»±―ΣΗΈ‘ύ‘λ≥…Υπ…ΥΘ§Εχ«“Ε‘»Ϊ…μΤδΥϋ‘ύΤς‘λ≥…“ΜΕ®ΒΡ…ΥΚΠΘΜ‘Ύ’β“Μ≤ΓάμΙΐ≥Χ÷–Θ§œΏΝΘΧεΕ‘œΗΑϊΡΎΗΤΒΡΒςΫΎΓΔ―θΉ‘”…Μυ–Έ≥…Β»ΨΏ”–Ζ«≥Θ÷Ί“ΣΒΡΉς”ΟΓΘ»γΚΈΖά÷ΈΗΈ‘ύ»±―Σ‘ΌΙύΉΔ ±œΏΝΘΧεΥπ…ΥΫΪ «ΫώΚσ Β―ι―–ΨΩΒΡ÷ΊΒψΓΘ
ΙΊΦϋ¥ ΘΚΗΈ‘ύΦ≤≤ΓΘΜœΏΝΘΧεΘΜ »±―Σ-‘ΌΙύΉΔΥπ…Υ
÷–ΆΦΖ÷άύΚ≈ΘΚR 575 ΈΡœΉ±ξ Ε¬κΘΚA ΈΡ’¬±ύΚ≈ΘΚ1007-8622(1999)04-0295-03
, http://www.100md.com
ΗΈ‘ύ «ΧεΡΎΉν¥σΒΡ¥ζ–ΜΤςΙΌΘ§œΏΝΘΧε «œΗΑϊ¥ζ–ΜΒΡ÷––Ρ≥ΓΥυΘ§œΏΝΘΧεΒΡ’ΐ≥ΘΚτΈϋΙΠΡήΚΆATP…ζ≥…Ε‘Ήι÷·ΤςΙΌΙΠΡήΦΑœΗΑϊΫαΙΙΒΡΆξ’ϊ–‘÷ΝΙΊ÷Ί“ΣΓΘΒΪœΏΝΘΧεΕ‘»±―ΣΓΔ»±―θΖ«≥ΘΟτΗ–Θ§»±―ΣΓΔ»±―θΒΡΗΈΉι÷·‘Ύ―ΣΝςΜ÷Η¥ΜρΗ¥―θΚσΤδΥπ…Υ≥ΧΕ»≤ΜΒΪΟΜ”–ΗΡ…ΤΘ§Ζ¥ΕχΫχ“Μ≤ΫΦ”÷ΊΘ§’β”κœΗΑϊΗΤ≥§‘ΊΓΔ―θΉ‘”…ΜυΒ»“ρΥΊ”–÷±Ϋ”ΒΡΙΊœΒ[1]ΓΘœΏΝΘΧε «ΝΣœΒΗΤ≥§‘ΊΓΔ―θΉ‘”…Μυ–Έ≥…ΚΆœΗΑϊΥάΆωΒΡ÷––ΡΜΖΫΎΓΘΫϋΡξΕ‘œΏΝΘΧε‘ΎΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡΉς”Ο―–ΨΩΫœΕύΘ§Ϋχ’ΙΫœΩλΘ§±ΨΈΡΨΆ’βΖΫΟφΒΡΡΎ»ίΉω“ΜΉέ ωΓΘ
1 œΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ”κ»±―Σ‘ΌΙύΉΔΗΤ≥§‘ΊΒΡΝΣœΒ
ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΩ…÷¬œΗΑϊΗΤ≥§‘ΊΘ§ «“ΐΤπΗΈœΗΑϊΥπ…ΥΒΡ‘≠“ρ÷°“ΜΘ§ΕχœΏΝΘΧεΨΏ”–«Ω¥σΒΡΨέΜΐCa2+ΡήΝΠΘ§Ε‘ΗΤάκΉ”Έ»Χ§ΤπΒΫ“ΜΕ®ΒΡΒςΫΎΉς”ΟΘ§ΒΪœΏΝΘΧε…ψCa2+ΡήΝΠ”–œόΘ§»τΥπ…Υ“ρΥΊ≤ΜΡή«ε≥ΐΘ§Ψ≠Ης÷÷ΆΨΨΕΉν÷’Ω…÷¬œΗΑϊΡΎΗΤ≥§‘Ί[2Θ§3]ΓΘ
ΫϋΡξΕ‘œΗΑϊΜΒΥάΒΡ―–ΨΩ±μΟςΘ§œΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ(Mitochondrial membrane permeability transition,MMPT)‘ΎœΗΑϊΥάΆω÷–ΤπΉ≈ΙΊΦϋΒΡΉς”Ο[4]ΓΘ‘ΎΗς÷÷Υπ…Υ“ρΥΊΒΡΉς”Οœ¬Θ§MMPT“ΜΒ©ΖΔ…ζΘ§œΏΝΘΧεΡΛΆ®ΆΗ–‘―ΗΥΌ…ΐΗΏΘ§œΗΑϊΥάΆωΥφ÷°ΖΔ…ζΓΘ“÷÷ΤMMPTΖΔ…ζΩ…―”ΜΚœΗΑϊΥάΆω[5]ΓΘΡΩ«Α»œΈΣMMPTΒΡΖΔ…ζΜζ÷ΤΑϋά®œΏΝΘΧεΡΎΡΛΒΡΖ«ΧΊ“λ–‘Υπ…ΥΚΆ“Μ÷÷ΨόΆ®Βά(Megapore)ΒΡΩΣΖ≈[4Θ§6]ΓΘΕχœΏΝΘΧεΡΎΡΛΖ«ΧΊ“λ–‘Υπ…ΥΩ…”…ΗΤ≥§‘Ί¥ΞΖΔΒΡ―θΉ‘”…ΜυΥπ…ΥΒ»‘λ≥…ΓΘΙΊ”ΎΒΦ÷¬MMPTΒΡΧΊ βΜζ÷ΤΒΡ―–ΨΩ±μΟςΘΚœΏΝΘΧεΜυ≤ψ÷–Ca2ΓFΒΡ¥σΝΩΨέΜΐ’ΐ «ΨόΆ®Βά(Megapore)ΩΣΖ≈ΒΡ‘≠“ρ÷°“Μ[5Θ§6]ΓΘΙ œΏΝΘΧεΗΤ≥§‘ΊΆ®ΙΐMMPTΩ…ΒΦ÷¬œΗΑϊ÷¬Υά–‘Υπ…ΥΓΘ
, ΑΌΡ¥“Ϋ“©
Ε‘ΨόΆ®ΒάΫχ“Μ≤Ϋ―–ΨΩΖΔœ÷Θ§œΏΝΘΧεΡΛ…œΒΡΨόΆ®ΒάΨΆ «÷±ΨΕΈΣ20(2 nm)ΒΡCa2+“άάΒ–ΓΩΉΘ§ΗΟ–ΓΩΉΨΏ”–―Γ‘ώ–‘Ά®ΆΗΉς”Ο[7]ΓΘ“―ΦχΕ®≥ωΗΟ–ΓΩΉΈΣœΏΝΘΧεΡΎΡΛΒΡΗΏΒγΒΦ–‘ΓΔœύΒ±ΒΆάκΉ”ΧΊ“λ–‘¥σΆ®ΒάΓΘœΏΝΘΧεΡΎCa2+ΚΆPiΒΡ…ΐΗΏΜρΜν–‘―θ¥ΧΦΛ ±–ΓΩΉΦ¥ΩΣΖ≈[6]Θ§Β±œΏΝΘΧεCa2+±Μ»Ξ≥ΐΚσ–ΓΩΉ―ΗΥΌΙΊ±’[4]ΘΜ…ζάμΦΝΝΩΒΡATP(>1mM)Ρή“÷÷ΤΤδΩΣΖ≈ΓΘ’ΐ≥Θ«ιΩωœ¬Θ§–ΓΩΉ≥…ΖίΜΖ«ΉΚΆΥΊ(cyclophilin)Έ§≥÷ΤδΩΣΖ≈ΓΘΜΖφΏΥΊA(cyclosporin A,CSA)Ρή”κΜΖ«ΉΚœΥΊΫαΚœ Ι–ΓΩΉΙΊ±’Θ§Ά§ ±Θ§CSAΨΏ”––ό≤ΙœΏΝΘΧεΡΎΡΛΒΡΙΠΡή(patch-clamp technique)[5,7].¥ΥΆβΘ§–ΓΩΉΒΡΩΣΖ≈ΜΙ”κœΏΝΘΧεΜυ÷ ΒΡpH”–ΙΊΘ§Β±œΏΝΘΡΎpH≥ Υα–‘ ±–ΓΩΉΒΡΩΣΖ≈Ιΐ≥Χ±δ¬ΐΓΘΝμΆβΘ§œΏΝΘΧεΆ®ΆΗ–‘ΗΡ±δΩ…”…œΏΝΘΧεΡΛΝΉ÷§ΒΡ»ΞθΘΜ·-‘ΌθΘΜ·―≠ΜΖΈ…¬““ΐΤπΓΘ
, ΑΌΡ¥“Ϋ“©
ΒΪΝμ”–―–ΨΩ»œΈΣΘ§‘ΎœΗΑϊΥπ…Υ÷–Θ§œΏΝΘΧε…ψΗΤ‘ωΕύ“≤–μ «ΦΧΖΔ–‘ΗΡ±δΒΡΫαΙϊΘ§ΕχœΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ(MMPT)≤≈ « ΦΕ·“ρΥΊΘ§“ρΕχΕ‘ΑϊΫ§ΗΤ≥§‘Ί‘ΎΉι÷·»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡ÷Ί“Σ–‘Χα≥ω÷ “…Θ§[8]Θ§“ρΈΣ‘Ύ¥σ σΗΈ‘ύ»±―θ35 min‘ΌΙύΉΔ10 minΒΡΡΘ–Ά÷–Θ§ΗΈΉι÷·ΒγΨΒœ¬Ά§“Μ ”“ΑΒΡΝΫΗωΗΈœΗΑϊ÷–Θ§“ΜΗωœΏΝΘΧε’ΐ≥ΘΘ§ΕχΝμ“ΜΗω‘ρΥ°÷Ή±δ–‘Θ§Κσ’Ώ“≤–μ“―”–œΏΝΘΧεΆ®ΆΗ–‘ΗΡ±δΘΜΕχ÷ΒΒΟΉΔ“βΒΡ «¥Υ ±»‘Έ¥ΦϊΤδΥϋœΗΑϊΤς”––ΈΧ§―ßΖΫΟφΒΡ±δΜ·[11]ΓΘ’βΫω «≥θ≤ΫΙέ≤λΒΡΫαΙϊΘ§“Σ≤ϊΟςΤδœξœΗΜζ÷ΤΘ§ΜΙ”–¥ΐΗϋΫχ“Μ≤Ϋ―–ΨΩΓΘ
2 œΏΝΘΧε‘Ύ―θΉ‘”…Μυ–Έ≥…÷–ΒΡΉς”Ο
ΜΤύ―Ώ ―θΜ·ΟΗΚΆ÷––‘ΑΉœΗΑϊ «―θΉ‘”…ΜυΒΡΝΫΗω÷Ί“Σά¥‘¥Θ§[9]Θ§»±―ΣΚΆ‘ΌΙύΉΔΨυΩ…≤ζ…ζΫœΕύ≥§―θΜ·ΈοΘ§“ΐΤπΉι÷·Υπ…ΥΓΘœΏΝΘΧε «ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΤΎΦδΜν–‘―θ≤ζ…ζΒΡ÷Ί“ΣΤςΙΌ[10]ΓΘ
2.1 ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΜν–‘―θ ΆΖ≈ΧΊΒψ”κΗΈœΗΑϊΥπ…ΥΒΡΙΊœΒΘΚΕ‘ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ―–ΨΩ±μΟςΘ§Η¥―θΚσΜν–‘―θ ΆΖ≈ ±œύ”κΗΈœΗΑϊΥπ…ΥΟή«–œύΙΊΓΘJaeschkeΒ»[11]ΉηΕœ≤ΩΖ÷ΗΈ‘ύ―ΣΝς45 minΚσ‘ΌΙύΉΔ(RP)≤ΜΆ§ ±ΦδΘ§ΖΔœ÷ΗΈΉι÷·≥ωœ÷ΝΥΥΪœύΥπ…ΥΓΘ‘ΎΜ÷Η¥ΙύΉΔΒΡ1 hΡΎΘ§―ΣΫ§ΉΣΑ±ΟΗΜν–‘…ΐΗΏΘ§¥ΥΈΣΗΈ‘ύ»±―Σ‘ΌΙύΉΔΒΡΔώœύΥπ…Υ(initial injury)ΘΜΫ”œ¬»ΞΒΡ5 h Θ§ΉΣΑ±ΟΗΒΡΜν–‘Έ§≥÷‘ΎΔώœύΥπ…ΥΥ°ΤΫΕχ‘ΎRPΚσ6ΓΪ24 hΘ§ΉΣΑ±ΟΗΜν–‘‘Ό¥Έœ‘÷χ…ΐΗΏΘ§±μΟς»±―Σ‘ΌΙύΉΔΗΈ‘ύΜΙ¥φ‘ΎΔρœύΥπ…Υ(progressive injury)ΓΘ¥”ΕχΩ…“‘Ω¥≥ωΗ¥―θ ±ΗΈœΗΑϊΥπ…Υ”κΜν–‘―θΒΡ ΆΖ≈ ±œύΨΏ”–÷±Ϋ”ΝΣœΒΓΘ Β―ιΜΙ÷ΛΟςRPΗΈ‘ύΒΡΔώœύΥπ…Υ”κΗΈώΦΩίΖώ œœΗΑϊ(KC)”–ΙΊΓΘKCΩ…“ρΗΤ≥§‘ΊΓΔΆΧ …Ής”ΟΜρ≤ΙΧεœΒΆ≥ΕχΦΛΜνΘ§ΜνΜ·ΒΡKC≥ΐ÷ς“Σ ΆΖ≈Μν–‘―θΆβΘ§ΜΙΩ… ΆΖ≈ΒΑΑΉΟΗΦΑΗς÷÷œΗΑϊ“ρΉ”Θ§ΫιΒΦΕ‘ΗΈœΗΑϊΒΡΕΨ–‘Ής”ΟΘ§ΒΦ÷¬―œ÷ΊΒΡΗΈ‘ύ‘ΌΙύΉΔΥπ…ΥΘΜΗ…»≈KCΙΠΡήΒΡ“©ΈοΨΏ”–Οςœ‘ΒΡ÷ΈΝΤΉς”Ο[12Θ§13]Θ§ΕχΔρœύΥπ…Υ”κΑΉœΗΑϊ‘ΎΨ÷≤ΩΒΡΨέΦ·ΒΫ“ΜΕ®ΝΩ≤ΔΦΛΜνΚσ ΆΖ≈ΕΨ–‘Ϋι÷ ”–ΙΊΘ§ΨΓΙήΑΉœΗΑϊ‘ΎRP≥θΤΎ‘ΎΗΈώΦ”κΡΎΤΛœΗΑϊ’≥ΗΫ‘ω«Ω≤ΔΩΣ ΦΨέΦ·Θ§ΒΪ≤Δ≤Μ≤Έ”κΗΈΉι÷·ΔώœύΥπ…Υ[13]ΓΘΆ§ ±ΜΙΖΔœ÷KCΒΡΔώœύΥπ…ΥΩ…ΦΛΜν÷––‘ΑΉœΗΑϊ(PMN)Θ§ ΙΤδ ΆΖ≈Μν–‘―θΦΑΒΑΑΉΟΗΒ»ΕΨ–‘Ϋι÷ ΙΞΜςΗΈœΗΑϊΘ§ΜΒΥάΗΈœΗΑϊΖ¥Ιΐά¥”÷Ω…¥ΌΫχ≤ΙΧεΦΛΜ·ΚΆPMNΜνΜ·Θ§»γ¥ΥΕώ–‘―≠ΜΖΘ§Ήν÷’ΒΦ÷¬―œ÷ΊΗΈœΗΑϊΥπ…ΥΓΘΒΞΩΥ¬ΓΩΙΧεΩ…Φθ…ΌPMN’≥ΗΫ≤Δ Ι‘ΌΙύΉΔΥπ…ΥΦθ«αΓΘ[14]ΓΘ
, http://www.100md.com
2.2 Μν–‘―θ”κ»±―Σ‘ΌΙύΉΔΚσΥπ…ΥΗΈœΗΑϊ‘ΎΗΈœΌ≈ί÷–ΒΡΖ÷≤ΦΘΚ“‘«Α―–ΨΩΖΔœ÷Θ§ΗΈ“Ε«–≥ΐΚσΗΈœΗΑϊΒΡ‘Ό…ζ÷ς“ΣΖΔ…ζ‘ΎΗΈ≈ίΔώ¥χΘ§ΕχΫα‘ζΕ·ΈοΗΈ―ΣΙή ΙΗΈ‘ύ»±―θΜρ”Σ―χ≤ΜΝΦ–‘‘γΤΎΗΈ”≤Μ·ΥυΖΔ…ζΒΡΗΈΥπΚΠ÷ς“ΣΖΔ…ζ‘ΎΔσ¥χΘ§’βΩ…Ϋβ ΆΈΣΔώ¥χΗΈœΗΑϊΩΩΫϋ÷–÷α―ΣΙήΘ§”≈œ»ΜώΒΟΗΜ”Ύ”Σ―χΒΡΈο÷ ΚΆΚ§―θ≥δΉψΒΡ―Σ“ΚΘ§“ρ¥Υ”≈œ»‘Ό…ζΘ§ΕχΔσ¥χΉι÷·PO2ΉνΒΆΘ§Ι Ήν»ί“Ή≥ωœ÷»±―θ–‘ΗΈΥπΚΠΚΆΗΈœΗΑϊΥάΆωΓΘ
Ρ«Ο¥Θ§ΗΈ‘ύ»±―ΣΚσ‘ΌΙύΉΔΥπ…ΥΒΡΧΊΒψ « ≤Ο¥ΡΊ?FlreemanΒ»ΒΡ―–ΨΩ»œΈΣΘΚΜΙ‘≠–ΆΙ»κΉΗ κΡ(GSH)‘ΎΗΈ‘ύ «÷Ί“ΣΒΡΉ‘”…Μυ«ε≥ΐΦΝΓΘ―–ΨΩΖΔœ÷Θ§¥σ σΗΈ‘ύ»±―Σ90 minΚσ‘ΌΙύΉΔΘ§ΗΈΉι÷·GSHΒΆ’Ώ(»γΫϊ ≥)ΗΈ‘ύΥπΚΠ≥ΧΕ»ΫœGSHΗΏ’ΏΗϋΈΣ―œ÷ΊΘ§≤Δ«“ΥπΚΠΗΈœΗΑϊ÷ς“ΣΖ÷≤Φ‘Ύ–Γ“ΕΦδΟ≈Ψ≤¬ω÷ήΈß«χ”ρ(Φ¥Δώ¥χ)Θ§’β÷÷±μœ÷‘ΎGSHΆξ»ΪœϊΚΡ’ΏΗϋΦ”Οςœ‘Θ§“ρΕχ÷ß≥÷―θΉ‘”…Μυ“ΐΤπΗΈΥπ…Υ’β“Μ¬έΕœ[15]ΓΘΆ§ ± Β―ι“≤÷ΛΟςΘ§”Ο¬»Μ·νή¥ΠάμΫϊ ≥ΒΡΕ·ΈοΘ§“‘‘ωΦ”GSH‘ΎΗΈΉι÷·ΒΡΚ§ΝΩΘ§ΫαΙϊ»±―ΣΚσΗΈœΗΑϊΜΒΥά≥ΧΕ»Οςœ‘Φθ«α[15]ΘΜ’β÷÷Ε‘»±―Σ‘ΌΙύΉΔΥπ…ΥΒΡ«χ¥χ–‘≤ν“λΘ§Ω…Ρή”κ“‘œ¬“ρΥΊ”–ΙΊΘΚΔΌ”κKupfferœΗΑϊ‘ΎΗΈώΦΒΡ≤ΜΨυ÷ –‘Ζ÷≤Φ”–ΙΊΓΘ¥σ σΗΈKupfferœΗΑϊ‘Φ’ΦΗΈ Β÷ ΒΡ2.1%Θ§ΒΪ“‘ΗΈ–Γ“Ε÷ή±Ώ¥χΖ÷≤ΦΫœΕύΓΘkupfferœΗΑϊΒΡΜνΜ·‘ΎΜζΧεΖάΈάΜζΡή÷–”–ΜΐΦΪΒΡΉς”ΟΘ§ΒΪΜνΜ·ΚσΒΡKupfferœΗΑϊ“‘ΤδΖ÷ΟΎΒΡΗς÷÷œΗΑϊ“ρΉ”(cytotines)Ής”Ο”ΎΗΈœΗΑϊΓΔΝΎΫϋΉι÷·ΦΑ»Ϊ…μΓΘ»±―ΣΓΔ‘ΌΙύΉΔΩ…÷±Ϋ”ΜρΦδΫ”ΦΛΜνKupfferœΗΑϊ(Μρ÷––‘ΑΉœΗΑϊ)Θ§Ω… ΆΖ≈Εύ÷÷œΗΑϊ“ρΉ”ΚΆΫι÷ [16]Θ§Τδ÷–Αϋά®O2¥ζ–Μ÷–Φδ≤ζΈοH2OΓΔO?2ΓΔOH?Ή‘”…ΜυΓΔO2ΦΑTNFΒ»ΓΘΔΎ”κ»±―ΣΚσΓΑΈόΙύΉΔΓ±(no-reflow)ΜρΓΑΈό―Σ‘ΌΙύΉΔΓ±(reflow-paradox)“ΐΤπΈΔ―≠ΜΖΈ…¬“”–ΙΊ [17]ΓΘΗΈώΦ «ΧΊΜ·ΒΡΟΪœΗ―ΣΙήΘ§Τδ÷±ΨΕ”κΤδΥϋΟΪœΗ―ΣΙήœύΥΤΘ§ΒΪΟΩΗωΗΈœΗΑϊΫτΝΎΝΫ…œΗΈώΦΘ§“ρΕχΕ‘ΟΪœΗ―ΣΙήΉη»ϊΒΡΡΆ ήΫœ«ΩΘ§‘Ύ…ζάμ«ιΩωœ¬“≤≥ΘΖΔ…ζΒΞΗω―ΣώΦΒΡΉη»ϊΓΘΒΪΗΈ‘ύ≥Λ ±Φδ»±―ΣΚσ”…”ΎΡΎΤΛΥ°÷ΉΓΔ―ΣώΦΡΎ―Σ“Κ≈®ΥθΦΑΈΔΨ≤¬ωΉη»ϊΒ»Θ§ Ι‘ΌΙύΉΔΚσ»±―Σ»±―θ»‘‘ΎΦΧ–χ(no-reflow)Θ§ΒΦ÷¬‘ΌΙύΉΔ≤Μ‘»Θ§Εχ‘ΌΙύΉΔΚσΓΑΈό―Σ‘ΌΙύΉΔΓ±(reflow-paradox) «”…”ΎΗ¥ΝςΜρΗ¥―θΦΛΜνΝΥΑΉœΗΑϊΘ§Ψ≠œΗΑϊ±μΟφ’≥ΗΫΖ÷Ή”ICAM-1ΓΔCD11/CD18ΓΔLΓΔP-―Γ‘ώΥΊ”κΡΎΤΛœΗΑϊάΈΙΧ’≥ΗΫΕχ“ΐΤπΗΈώΦΉη»ϊΓΘΆ®Ιΐ“‘…œΝΫ÷÷ΖΫ ΫΘ§ ΙΗ¥ΝςΚσΜν–‘―θ ΆΖ≈‘ωΕύ≤Δœό÷Τ‘ΎΨ÷≤ΩΘ§Ά§ ±Θ§”…”Ύ―θΉ‘”…ΜυΤΫΨυ ΌΟϋΚήΕΧΘ§Ϋω10?3Οκ[18]Θ§‘ΎΈΔ―≠ΜΖΈ…¬“ΒΡ«ιΩωœ¬≤ΜΡήΝΔΦ¥‘ΎΗΈΉι÷·≤Ξ…ΔΜρ¥σΝΩ ΆΖ≈»κ―Σ―≠÷–Θ§‘λ≥…Ψ÷≤ΩΉι÷·―œ÷ΊΥπ…Υ(»γΟ≈Ιή«χ)Θ§Rymsa―–ΨΩΖΔœ÷Θ§RP≥θΤΎ‘ΎΟ≈¬ω÷ήΈß«χ”ρΖ÷άκ≥ωΨΏ”–Κή«ΩΆΧ …Μν–‘ΒΡ¥σKCΘ§ΤδΉ‘ΖΔ≤ζ…ζΒΡO?2‘ωΦ”3±Ε“‘…œΓΘSuzukiΒ»‘ΎΗΈ‘ύ»±―Σ90 min‘ΌΙύ60 minΒΡΉι÷·―ßΙέ≤λ÷–ΖΔœ÷Θ§≤Γ±δ÷ς“ΣΖ÷≤Φ‘ΎΗΈ≈ίΔώ¥χ÷ΝΔρ¥χΘ§Ω…ΦϊΗΈœΗΑϊ―œ÷Ί±δ–‘Ή«÷ΉΓΔΗΈώΦπω―Σ[9]ΓΘΙΊ”ΎΤδΥπ…ΥΖ÷≤Φ≤ΩΈΜΒΡ»Ζ«–Μζ÷ΤΜΙ”–¥ΐΫχ“Μ≤Ϋ―–ΨΩΓΘ
, ΑΌΡ¥“Ϋ“©
ΒΪGonzalez-Flecha[15]―–ΨΩΖΔœ÷Θ§ΗΈœΌ≈ίΔρΓΔΔσ¥χΕ‘»±―Σ‘ΌΙύΉΔΥπ…Υ±»ΫœΟτΗ–Θ§ΡΩ«Α―–ΨΩ»œΈΣΗΈ≈ίΈό¬έ‘ΎœΗΑϊΤςΫαΙΙΜρΟΗΒΡΉιΖίΕΦ”–Υυ≤ΜΆ§Θ§’β «ΥϋΕ‘»±―Σ‘ΌΙύΉΔΥπ…ΥΟτΗ––‘≤ΜΆ§ΒΡΜυ¥ΓΓΘΡΩ«ΑΕ‘ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…ΥΟτΗ––‘ΒΡΉι÷·―ßΕ®ΈΜ»‘”–’υ“ιΘ§ΜΙ”–¥ΐΫχ“Μ≤ΫΧΫΧ÷ΓΘ
3 »±―Σ‘ΌΙύΉΔœΏΝΘΧεΚτΈϋΜν–‘ΒΡ±δΜ·
«ΑΟφ“―Ψ≠ΧαΒΫœΏΝΘΧε‘ΎœΗΑϊΗΤ≥§‘ΊΦΑ―θΉ‘”…Μυ–Έ≥…÷–ΒΡΒςΫΎΉς”ΟΓΘΒΪΒ±’β÷÷ΒςΫΎΉς”Ο≥§Ιΐ“ΜΕ®œόΕ»Θ§±Ί»Μ“ΐΤπœΏΝΘΧεΉ‘…μΥπΚΠΘ§’β÷÷ΥπΚΠ≥ωœ÷«Α Ήœ»≥ωœ÷“ΜœΒΝ–¥ζ≥Ξ–‘ΗΡ±δΘ§Ι ΝΥΫβΤδ±δΜ·Ιφ¬…Ε‘»±―Σ‘ΌΙύΉΔΥπ…ΥΒΡΖά÷ΈΨΏ”–ΜΐΦΪΒΡΉς”ΟΓΘ
ΤάΦέœΏΝΘΧεΚτΈϋΜν–‘≥Θ”ΟΚτΈϋΩΊ÷Τ¬ (RCR)ΚΆΝΉ―θ±»(P/O)ά¥±μ ΨΓΘΥϋΟ« «Ζ¥”≥œΏΝΘΧεΆξ’ϊ–‘ΓΔ―θΜ·ΝΉΥαΜ·≈ΦΝΣ≥ΧΕ»ΦΑ―θΜ·ΝΉΥαΜ·–߬ ΒΡΝιΟτ÷Η±ξΓΘΗΈ‘ύ»±―Σ‘ΌΙύΉΔ ±œΏΝΘΧεΆ®ΆΗ–‘ΒΡΗΡ±δΓΔœΏΝΘΧεΡΎΟΗΚΆΗ®ΟΗΒΡΕΣ ßΘ§“‘ΦΑ”…¥Υ‘λ≥…ΒΡ―θΜ·ΝΉΥαΜ·’œΑ≠Θ§ΨυΩ…”ΑœλœΏΝΘΧεΚτΈϋΜν–‘ΓΘIsozaki‘Ύ―–ΨΩΦδΕœΦΑ≥÷–χΗΈ‘ύ»±―ΣΕ‘œΏΝΘΧεΙΠΡήΒΡ”ΑœλΚσΖΔœ÷Θ§ΗΈ‘ύ‘Ύ≥÷–χ»±―Σ30 minΚσΕ‘RCRΦΑP/O”Αœλ≤Μ¥σΘΜ»τΫΪ»±―ΣΉή ±Φδ…ηΕ®‘Ύ60 minΘ§Ψ≠Ιέ≤λΖΔœ÷ΦδΕœ»±―Σ’ΏœΏΝΘΧεΙΠΡήΟςœ‘ΚΟ”Ύ≥÷–χ»±―Σ’ΏΘ§±μΟςΥφ≥÷–χ»±―Σ ±ΦδΒΡ―”≥ΛΘ§œΏΝΘΧεΒΡΆξ’ϊ–‘ ήΒΫ“ΜΕ®ΒΡ”ΑœλΓΘ
, ΑΌΡ¥“Ϋ“©
KorokawaΒ»―–ΨΩΗΈ¥σ≤ΩΖ÷«–≥ΐΚσ≤–ΗΈœΗΑϊœΏΝΘΧεΒΡΙΠΡή ±ΦϊΒΫΘ§ ÷ θΚσΒΎ“ΜΧλCRCΦΑR3Ϋœ ÷ θ«ΑΟςœ‘‘ωΗΏΘ§2ΓΪ5 dΚσ÷πΫΞΫΒΒΆ÷Ν θ«ΑΥ°ΤΫΘ§ΥΒΟς≤–ΗΈ¥ζ–ΜΜν‘ΨœΏΝΘΧεΙΠΡή¥ζ≥Ξ–‘‘ω«ΩΓΘΒΪKobayashi[19]Ε‘»±―Σ‘ΌΙύΉΔ ±ΗΈ‘ύœΏΝΘΧεΚτΈϋΜν–‘Ιέ≤λΖΔœ÷Θ§»±―Σ120 minœΏΝΘΧεRCRΓΔR3Ϋœ»±―Σ«ΑΟςœ‘ΫΒΒΆΘ§‘ΌΙύΉΔΚσΥδ”–ΥυΧαΗΏΘ§ΒΪ»‘Οςœ‘ΒΆ”Ύ θ«ΑΥ°ΤΫΓΘGonzalez-Flecha[10]ΒΡ―–ΨΩ”κ“‘…œœύΥΤΘ§Ιέ≤λΝΥ»±―Σ60 minΓΔ120 minΦΑ180 minΦΑ‘ΌΙύΉΔ30 min ±œΏΝΘΧεΚτΈϋΙΠΡήΒΡΗΡ±δΘ§Φϊ»±―ΣΚσRCRΓΔR3Ψυ≤ΜΆ§≥ΧΕ»ΫΒΒΆΘ§≤ΔΥφ»±―Σ ±Φδ―”≥ΛΘ§’β÷÷±δΜ·‘ΫΟςœ‘Θ§R4‘ρΈόΟςœ‘±δΜ·ΘΜ‘ΌΙύΉΔ30 min ±Θ§“‘…œ÷Η±ξ≤ΜΒΪΟΜ”–…ΐΗΏΖ¥ΕχΫœ»±―ΣΚσΟςœ‘ΫΒΒΆΘ§ΒΪ»±―Σ180 min‘ΌΙύΉΔ30 minΉι‘ρ±δΜ·≤ΜΟςœ‘ΜρΈό±δΜ·Θ§“≤–μ¥Υ ±œΏΝΘΧε¥ζ≥Ξ“―ΒΫΦΪœόΘ§Ε‘Ης÷÷¥ΧΦΛΟτΗ––‘ΫΒΒΆΥυ÷¬ΓΘ
Ής’ΏΦρΫιΘΚΚΈ–ßΕΪ(1962-)Θ§Ρ–Θ§ΥΕ ΩΘ§÷ς÷Έ“Ϋ Π
, http://www.100md.com
Ής’ΏΒΞΈΜΘΚΚΈ–ßΕΪΘΚΫβΖ≈ΨϋΒΎ“Μ“Ϋ‘ΚΗΈΒ®ΆβΩΤ÷––ΡΘ§άΦ÷ί730030
Ε≠Φ“Κη ≤ΧΨΑ–όΘΚΒΎ»ΐΨϋ“Ϋ¥σ―ßΈςΡœ“Ϋ‘ΚΗΈΒ®ΆβΩΤ÷––ΡΘ§÷Ί«λ 400038
≤ΈΩΦΈΡœΉΘΚ
[1]liu W,Schob O, Pugmire JE,et al.Glycohydrolases as markers of hepatic ischemia-reperfusion injury and recovery [J].Hepatology,1996,24:157.
[2]Hatanaka N,Kamijke W,Shimizu S,et al.Ca2ΓFrelease from mitochondrial induces cytosiolic enzyme leakage in anoxis liver [J].J Surg Res,1995,58:485.
, ΑΌΡ¥“Ϋ“©
[3]Masini A,Gallesi D,Giovannini F,et al. Membrane potential of hepatic mitochondrial after cute cocaine administration in rat-the role of mitochondrial reduced glutathione [J].Hepatology,1997,25:305.
[4]Saris N-E L,Eriksson KD.Mitochondrial dysfunction in ischemia-reperfusion [J].Acta Anaesthesiologica Scandinavia,1995,29:(suppl 107):171.
[5]Geexaerts MD,Ronveavx-Dupal MF,Lemasters JJ,et al.Cytosilic free Ca2+and proteolysis in lethal oxidative injury in endothelial cell[J].Am J Physiol,1991,261(5):C889.
, http://www.100md.com
[6]Szabo I, Zoratti M.The giant channel of the inner mitochondrial membrane is inhibited by cyclosporin A [J].J Biol,Chem,1991,266:3376.
[7]Bernard P,Vassanell S,Veronese P,et al.Modulation of the mitochondrial permeability transition pore[J].J Biol Chem,1992,267:2934.
[8]Jacobs WR,Sgambati M,Gomez G,et al.Role of cytosolic Ca 2+ in renal tubule damage induced by anoxia [J].Am J Physiol,1991,260:C545.
, ΑΌΡ¥“Ϋ“©
[9]Suzuki S,Toledo-pereyra LH,Rodriguez FL,et al. Neutrophil infilration as an important factor in liver ischemia and reperfusion injury[J].Transplantation,1993,55(6):1265.
[10]Gonzalez-Flecha B, Cutrin JC,Boveris A.Time course and mechanism of oxidative stress and tissue damage in rat liver subjected to in vivo ischemia-reperfusion[J]. J Clin Invest 1993,91:456.
[11]Jaeschke H,Farbood A,Smith W.Neutrophils contribute to ischemia/reperfusion injury in rat liver in vivo [J].FASEB J ,1990,4:3355.
, http://www.100md.com
[12]Brass CA,Roberts TG.Hepatic free radical production after cold storage;kupffer cell-dependent and independent mechanisms in rats [J].Gastroenterology,1995,108:1167.
[13]Niwano M,Arii S,Monden K,et al. Aneliovation of sinusoided endothelial cell damage by kupffer cell blockade during cell preservation of rat liver [J].J Surg Res,1997,72:36.
[14]Marubayashi S,Oshire Y,Maeda T,et al.Protective effect of monoclonal antibodies to adhesion molecules on rat liver ischemia-reperfusion injury [J].Surgery,1997,122:45.
, ΑΌΡ¥“Ϋ“©
[15]Jennische E. Possible influence of glutathine on postischemic liver injury [J].Acta Pathol Microbiol Scand,[C]1984,92:55.
[16]Shibuyo H,Ohkohchi N,Seya K,et al.Kupffer cell generate superoxide anions and modulate reperfusion injury in rat liver after cold preservation [J].Hepatology,1997,25:356.
[17]Menger MD.Microcirculatory disturbance secondary to ischemia-reperfusion [J].Transplantation,1995,27(5):2863.
[18]¬§γ≤ΤΦΘ§ Ζ≥¬»ΟΘ§Άτ ΚλΘ§Β».¥σ σΗΈ»±―Σ‘ΌΙύΝςΤΎΦδ÷§÷ Ιΐ―θΜ·Ή‘”…ΜυΕ·Χ§±δΜ·ΦΑ≤ΓάμΗΡ±δΆ§≤ΫΙέ≤λ.÷–ΜΣ Β―ιΆβΩΤ‘”÷ΨΘ§1992Θ§9ΘΚ11.
[19]Kobayashi H,Nonami T,Kurokawa T,et al.Mechanism and prevention of ischemia-reperfusion liver injury in rats [J] J Surg Res,1991,51:240.
’Ηε»’ΤΎΘΚ1999-01-20, ΑΌΡ¥“Ϋ“©
ΒΞΈΜΘΚΚΈ–ßΕΪΘΚΫβΖ≈ΨϋΒΎ“Μ“Ϋ‘ΚΗΈΒ®ΆβΩΤ÷––ΡΘ§άΦ÷ί730030 Ε≠Φ“Κη ≤ΧΨΑ–όΘΚΒΎ»ΐΨϋ“Ϋ¥σ―ßΈςΡœ“Ϋ‘ΚΗΈΒ®ΆβΩΤ÷––ΡΘ§÷Ί«λ 400038
ΙΊΦϋ¥ ΘΚΗΈ‘ύΦ≤≤ΓΘΜœΏΝΘΧεΘΜ»±―Σ-‘ΌΙύΉΔΥπ…Υ
œΏΝΘΧε‘ΎΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡΉς”ΟΚΈ–ßΕΪ Ήέ ω Ε≠Φ“Κη ≤ΧΨΑ–ό …σ–Θ
’Σ“ΣΘΚΗΈ‘ύΆβΩΤΓΔ–ίΩΥΒ»≤ΓάμΙΐ≥Χ÷–ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ≤ΜΩ…±ήΟβΘ§ΗΟΙΐ≥Χ“‘»±―ΣΓΔ»±―θΓΔΡήΝΩΙ©”Π÷–ΕœΈΣ ΦΕ·“ρΥΊΘ§Ψ≠œΗΑϊΗΤ≥§‘ΊΓΔ―θΉ‘”…Μυ–Έ≥…ΦΑΑΉœΗΑϊΫΰ»σ»ΐΗω÷Ί“ΣΜΖΫΎΘ§Ά®ΙΐœΗΑϊ“ρΉ”―Ή–‘Ϋι÷ ΒΡ ΆΖ≈Θ§≤ΜΒΪΕ‘»±―ΣΗΈ‘ύ‘λ≥…Υπ…ΥΘ§Εχ«“Ε‘»Ϊ…μΤδΥϋ‘ύΤς‘λ≥…“ΜΕ®ΒΡ…ΥΚΠΘΜ‘Ύ’β“Μ≤ΓάμΙΐ≥Χ÷–Θ§œΏΝΘΧεΕ‘œΗΑϊΡΎΗΤΒΡΒςΫΎΓΔ―θΉ‘”…Μυ–Έ≥…Β»ΨΏ”–Ζ«≥Θ÷Ί“ΣΒΡΉς”ΟΓΘ»γΚΈΖά÷ΈΗΈ‘ύ»±―Σ‘ΌΙύΉΔ ±œΏΝΘΧεΥπ…ΥΫΪ «ΫώΚσ Β―ι―–ΨΩΒΡ÷ΊΒψΓΘ
÷–ΆΦΖ÷άύΚ≈ΘΚR 575 ΈΡœΉ±ξ Ε¬κΘΚA ΈΡ’¬±ύΚ≈ΘΚ1007-8622(1999)04-0295-03
, http://www.100md.com
ΗΈ‘ύ «ΧεΡΎΉν¥σΒΡ¥ζ–ΜΤςΙΌΘ§œΏΝΘΧε «œΗΑϊ¥ζ–ΜΒΡ÷––Ρ≥ΓΥυΘ§œΏΝΘΧεΒΡ’ΐ≥ΘΚτΈϋΙΠΡήΚΆATP…ζ≥…Ε‘Ήι÷·ΤςΙΌΙΠΡήΦΑœΗΑϊΫαΙΙΒΡΆξ’ϊ–‘÷ΝΙΊ÷Ί“ΣΓΘΒΪœΏΝΘΧεΕ‘»±―ΣΓΔ»±―θΖ«≥ΘΟτΗ–Θ§»±―ΣΓΔ»±―θΒΡΗΈΉι÷·‘Ύ―ΣΝςΜ÷Η¥ΜρΗ¥―θΚσΤδΥπ…Υ≥ΧΕ»≤ΜΒΪΟΜ”–ΗΡ…ΤΘ§Ζ¥ΕχΫχ“Μ≤ΫΦ”÷ΊΘ§’β”κœΗΑϊΗΤ≥§‘ΊΓΔ―θΉ‘”…ΜυΒ»“ρΥΊ”–÷±Ϋ”ΒΡΙΊœΒ[1]ΓΘœΏΝΘΧε «ΝΣœΒΗΤ≥§‘ΊΓΔ―θΉ‘”…Μυ–Έ≥…ΚΆœΗΑϊΥάΆωΒΡ÷––ΡΜΖΫΎΓΘΫϋΡξΕ‘œΏΝΘΧε‘ΎΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡΉς”Ο―–ΨΩΫœΕύΘ§Ϋχ’ΙΫœΩλΘ§±ΨΈΡΨΆ’βΖΫΟφΒΡΡΎ»ίΉω“ΜΉέ ωΓΘ
1 œΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ”κ»±―Σ‘ΌΙύΉΔΗΤ≥§‘ΊΒΡΝΣœΒ
ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΩ…÷¬œΗΑϊΗΤ≥§‘ΊΘ§ «“ΐΤπΗΈœΗΑϊΥπ…ΥΒΡ‘≠“ρ÷°“ΜΘ§ΕχœΏΝΘΧεΨΏ”–«Ω¥σΒΡΨέΜΐCa2+ΡήΝΠΘ§Ε‘ΗΤάκΉ”Έ»Χ§ΤπΒΫ“ΜΕ®ΒΡΒςΫΎΉς”ΟΘ§ΒΪœΏΝΘΧε…ψCa2+ΡήΝΠ”–œόΘ§»τΥπ…Υ“ρΥΊ≤ΜΡή«ε≥ΐΘ§Ψ≠Ης÷÷ΆΨΨΕΉν÷’Ω…÷¬œΗΑϊΡΎΗΤ≥§‘Ί[2Θ§3]ΓΘ
ΫϋΡξΕ‘œΗΑϊΜΒΥάΒΡ―–ΨΩ±μΟςΘ§œΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ(Mitochondrial membrane permeability transition,MMPT)‘ΎœΗΑϊΥάΆω÷–ΤπΉ≈ΙΊΦϋΒΡΉς”Ο[4]ΓΘ‘ΎΗς÷÷Υπ…Υ“ρΥΊΒΡΉς”Οœ¬Θ§MMPT“ΜΒ©ΖΔ…ζΘ§œΏΝΘΧεΡΛΆ®ΆΗ–‘―ΗΥΌ…ΐΗΏΘ§œΗΑϊΥάΆωΥφ÷°ΖΔ…ζΓΘ“÷÷ΤMMPTΖΔ…ζΩ…―”ΜΚœΗΑϊΥάΆω[5]ΓΘΡΩ«Α»œΈΣMMPTΒΡΖΔ…ζΜζ÷ΤΑϋά®œΏΝΘΧεΡΎΡΛΒΡΖ«ΧΊ“λ–‘Υπ…ΥΚΆ“Μ÷÷ΨόΆ®Βά(Megapore)ΒΡΩΣΖ≈[4Θ§6]ΓΘΕχœΏΝΘΧεΡΎΡΛΖ«ΧΊ“λ–‘Υπ…ΥΩ…”…ΗΤ≥§‘Ί¥ΞΖΔΒΡ―θΉ‘”…ΜυΥπ…ΥΒ»‘λ≥…ΓΘΙΊ”ΎΒΦ÷¬MMPTΒΡΧΊ βΜζ÷ΤΒΡ―–ΨΩ±μΟςΘΚœΏΝΘΧεΜυ≤ψ÷–Ca2ΓFΒΡ¥σΝΩΨέΜΐ’ΐ «ΨόΆ®Βά(Megapore)ΩΣΖ≈ΒΡ‘≠“ρ÷°“Μ[5Θ§6]ΓΘΙ œΏΝΘΧεΗΤ≥§‘ΊΆ®ΙΐMMPTΩ…ΒΦ÷¬œΗΑϊ÷¬Υά–‘Υπ…ΥΓΘ
, http://www.100md.com
Ε‘ΨόΆ®ΒάΫχ“Μ≤Ϋ―–ΨΩΖΔœ÷Θ§œΏΝΘΧεΡΛ…œΒΡΨόΆ®ΒάΨΆ «÷±ΨΕΈΣ20
, http://www.100md.com
ΒΪΝμ”–―–ΨΩ»œΈΣΘ§‘ΎœΗΑϊΥπ…Υ÷–Θ§œΏΝΘΧε…ψΗΤ‘ωΕύ“≤–μ «ΦΧΖΔ–‘ΗΡ±δΒΡΫαΙϊΘ§ΕχœΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ(MMPT)≤≈ « ΦΕ·“ρΥΊΘ§“ρΕχΕ‘ΑϊΫ§ΗΤ≥§‘Ί‘ΎΉι÷·»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡ÷Ί“Σ–‘Χα≥ω÷ “…Θ§[8]Θ§“ρΈΣ‘Ύ¥σ σΗΈ‘ύ»±―θ35 min‘ΌΙύΉΔ10 minΒΡΡΘ–Ά÷–Θ§ΗΈΉι÷·ΒγΨΒœ¬Ά§“Μ ”“ΑΒΡΝΫΗωΗΈœΗΑϊ÷–Θ§“ΜΗωœΏΝΘΧε’ΐ≥ΘΘ§ΕχΝμ“ΜΗω‘ρΥ°÷Ή±δ–‘Θ§Κσ’Ώ“≤–μ“―”–œΏΝΘΧεΆ®ΆΗ–‘ΗΡ±δΘΜΕχ÷ΒΒΟΉΔ“βΒΡ «¥Υ ±»‘Έ¥ΦϊΤδΥϋœΗΑϊΤς”––ΈΧ§―ßΖΫΟφΒΡ±δΜ·[11]ΓΘ’βΫω «≥θ≤ΫΙέ≤λΒΡΫαΙϊΘ§“Σ≤ϊΟςΤδœξœΗΜζ÷ΤΘ§ΜΙ”–¥ΐΗϋΫχ“Μ≤Ϋ―–ΨΩΓΘ
2 œΏΝΘΧε‘Ύ―θΉ‘”…Μυ–Έ≥…÷–ΒΡΉς”Ο
ΜΤύ―Ώ ―θΜ·ΟΗΚΆ÷––‘ΑΉœΗΑϊ «―θΉ‘”…ΜυΒΡΝΫΗω÷Ί“Σά¥‘¥Θ§[9]Θ§»±―ΣΚΆ‘ΌΙύΉΔΨυΩ…≤ζ…ζΫœΕύ≥§―θΜ·ΈοΘ§“ΐΤπΉι÷·Υπ…ΥΓΘœΏΝΘΧε «ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΤΎΦδΜν–‘―θ≤ζ…ζΒΡ÷Ί“ΣΤςΙΌ[10]ΓΘ
2.1 ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΜν–‘―θ ΆΖ≈ΧΊΒψ”κΗΈœΗΑϊΥπ…ΥΒΡΙΊœΒΘΚΕ‘ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ―–ΨΩ±μΟςΘ§Η¥―θΚσΜν–‘―θ ΆΖ≈ ±œύ”κΗΈœΗΑϊΥπ…ΥΟή«–œύΙΊΓΘJaeschkeΒ»[11]ΉηΕœ≤ΩΖ÷ΗΈ‘ύ―ΣΝς45 minΚσ‘ΌΙύΉΔ(RP)≤ΜΆ§ ±ΦδΘ§ΖΔœ÷ΗΈΉι÷·≥ωœ÷ΝΥΥΪœύΥπ…ΥΓΘ‘ΎΜ÷Η¥ΙύΉΔΒΡ1 hΡΎΘ§―ΣΫ§ΉΣΑ±ΟΗΜν–‘…ΐΗΏΘ§¥ΥΈΣΗΈ‘ύ»±―Σ‘ΌΙύΉΔΒΡΔώœύΥπ…Υ(initial injury)ΘΜΫ”œ¬»ΞΒΡ5 h Θ§ΉΣΑ±ΟΗΒΡΜν–‘Έ§≥÷‘ΎΔώœύΥπ…ΥΥ°ΤΫΕχ‘ΎRPΚσ6ΓΪ24 hΘ§ΉΣΑ±ΟΗΜν–‘‘Ό¥Έœ‘÷χ…ΐΗΏΘ§±μΟς»±―Σ‘ΌΙύΉΔΗΈ‘ύΜΙ¥φ‘ΎΔρœύΥπ…Υ(progressive injury)ΓΘ¥”ΕχΩ…“‘Ω¥≥ωΗ¥―θ ±ΗΈœΗΑϊΥπ…Υ”κΜν–‘―θΒΡ ΆΖ≈ ±œύΨΏ”–÷±Ϋ”ΝΣœΒΓΘ Β―ιΜΙ÷ΛΟςRPΗΈ‘ύΒΡΔώœύΥπ…Υ”κΗΈώΦΩίΖώ œœΗΑϊ(KC)”–ΙΊΓΘKCΩ…“ρΗΤ≥§‘ΊΓΔΆΧ …Ής”ΟΜρ≤ΙΧεœΒΆ≥ΕχΦΛΜνΘ§ΜνΜ·ΒΡKC≥ΐ÷ς“Σ ΆΖ≈Μν–‘―θΆβΘ§ΜΙΩ… ΆΖ≈ΒΑΑΉΟΗΦΑΗς÷÷œΗΑϊ“ρΉ”Θ§ΫιΒΦΕ‘ΗΈœΗΑϊΒΡΕΨ–‘Ής”ΟΘ§ΒΦ÷¬―œ÷ΊΒΡΗΈ‘ύ‘ΌΙύΉΔΥπ…ΥΘΜΗ…»≈KCΙΠΡήΒΡ“©ΈοΨΏ”–Οςœ‘ΒΡ÷ΈΝΤΉς”Ο[12Θ§13]Θ§ΕχΔρœύΥπ…Υ”κΑΉœΗΑϊ‘ΎΨ÷≤ΩΒΡΨέΦ·ΒΫ“ΜΕ®ΝΩ≤ΔΦΛΜνΚσ ΆΖ≈ΕΨ–‘Ϋι÷ ”–ΙΊΘ§ΨΓΙήΑΉœΗΑϊ‘ΎRP≥θΤΎ‘ΎΗΈώΦ”κΡΎΤΛœΗΑϊ’≥ΗΫ‘ω«Ω≤ΔΩΣ ΦΨέΦ·Θ§ΒΪ≤Δ≤Μ≤Έ”κΗΈΉι÷·ΔώœύΥπ…Υ[13]ΓΘΆ§ ±ΜΙΖΔœ÷KCΒΡΔώœύΥπ…ΥΩ…ΦΛΜν÷––‘ΑΉœΗΑϊ(PMN)Θ§ ΙΤδ ΆΖ≈Μν–‘―θΦΑΒΑΑΉΟΗΒ»ΕΨ–‘Ϋι÷ ΙΞΜςΗΈœΗΑϊΘ§ΜΒΥάΗΈœΗΑϊΖ¥Ιΐά¥”÷Ω…¥ΌΫχ≤ΙΧεΦΛΜ·ΚΆPMNΜνΜ·Θ§»γ¥ΥΕώ–‘―≠ΜΖΘ§Ήν÷’ΒΦ÷¬―œ÷ΊΗΈœΗΑϊΥπ…ΥΓΘΒΞΩΥ¬ΓΩΙΧεΩ…Φθ…ΌPMN’≥ΗΫ≤Δ Ι‘ΌΙύΉΔΥπ…ΥΦθ«αΓΘ[14]ΓΘ
, http://www.100md.com
2.2 Μν–‘―θ”κ»±―Σ‘ΌΙύΉΔΚσΥπ…ΥΗΈœΗΑϊ‘ΎΗΈœΌ≈ί÷–ΒΡΖ÷≤ΦΘΚ“‘«Α―–ΨΩΖΔœ÷Θ§ΗΈ“Ε«–≥ΐΚσΗΈœΗΑϊΒΡ‘Ό…ζ÷ς“ΣΖΔ…ζ‘ΎΗΈ≈ίΔώ¥χΘ§ΕχΫα‘ζΕ·ΈοΗΈ―ΣΙή ΙΗΈ‘ύ»±―θΜρ”Σ―χ≤ΜΝΦ–‘‘γΤΎΗΈ”≤Μ·ΥυΖΔ…ζΒΡΗΈΥπΚΠ÷ς“ΣΖΔ…ζ‘ΎΔσ¥χΘ§’βΩ…Ϋβ ΆΈΣΔώ¥χΗΈœΗΑϊΩΩΫϋ÷–÷α―ΣΙήΘ§”≈œ»ΜώΒΟΗΜ”Ύ”Σ―χΒΡΈο÷ ΚΆΚ§―θ≥δΉψΒΡ―Σ“ΚΘ§“ρ¥Υ”≈œ»‘Ό…ζΘ§ΕχΔσ¥χΉι÷·PO2ΉνΒΆΘ§Ι Ήν»ί“Ή≥ωœ÷»±―θ–‘ΗΈΥπΚΠΚΆΗΈœΗΑϊΥάΆωΓΘ
Ρ«Ο¥Θ§ΗΈ‘ύ»±―ΣΚσ‘ΌΙύΉΔΥπ…ΥΒΡΧΊΒψ « ≤Ο¥ΡΊ?FlreemanΒ»ΒΡ―–ΨΩ»œΈΣΘΚΜΙ‘≠–ΆΙ»κΉΗ κΡ(GSH)‘ΎΗΈ‘ύ «÷Ί“ΣΒΡΉ‘”…Μυ«ε≥ΐΦΝΓΘ―–ΨΩΖΔœ÷Θ§¥σ σΗΈ‘ύ»±―Σ90 minΚσ‘ΌΙύΉΔΘ§ΗΈΉι÷·GSHΒΆ’Ώ(»γΫϊ ≥)ΗΈ‘ύΥπΚΠ≥ΧΕ»ΫœGSHΗΏ’ΏΗϋΈΣ―œ÷ΊΘ§≤Δ«“ΥπΚΠΗΈœΗΑϊ÷ς“ΣΖ÷≤Φ‘Ύ–Γ“ΕΦδΟ≈Ψ≤¬ω÷ήΈß«χ”ρ(Φ¥Δώ¥χ)Θ§’β÷÷±μœ÷‘ΎGSHΆξ»ΪœϊΚΡ’ΏΗϋΦ”Οςœ‘Θ§“ρΕχ÷ß≥÷―θΉ‘”…Μυ“ΐΤπΗΈΥπ…Υ’β“Μ¬έΕœ[15]ΓΘΆ§ ± Β―ι“≤÷ΛΟςΘ§”Ο¬»Μ·νή¥ΠάμΫϊ ≥ΒΡΕ·ΈοΘ§“‘‘ωΦ”GSH‘ΎΗΈΉι÷·ΒΡΚ§ΝΩΘ§ΫαΙϊ»±―ΣΚσΗΈœΗΑϊΜΒΥά≥ΧΕ»Οςœ‘Φθ«α[15]ΘΜ’β÷÷Ε‘»±―Σ‘ΌΙύΉΔΥπ…ΥΒΡ«χ¥χ–‘≤ν“λΘ§Ω…Ρή”κ“‘œ¬“ρΥΊ”–ΙΊΘΚΔΌ”κKupfferœΗΑϊ‘ΎΗΈώΦΒΡ≤ΜΨυ÷ –‘Ζ÷≤Φ”–ΙΊΓΘ¥σ σΗΈKupfferœΗΑϊ‘Φ’ΦΗΈ Β÷ ΒΡ2.1%Θ§ΒΪ“‘ΗΈ–Γ“Ε÷ή±Ώ¥χΖ÷≤ΦΫœΕύΓΘkupfferœΗΑϊΒΡΜνΜ·‘ΎΜζΧεΖάΈάΜζΡή÷–”–ΜΐΦΪΒΡΉς”ΟΘ§ΒΪΜνΜ·ΚσΒΡKupfferœΗΑϊ“‘ΤδΖ÷ΟΎΒΡΗς÷÷œΗΑϊ“ρΉ”(cytotines)Ής”Ο”ΎΗΈœΗΑϊΓΔΝΎΫϋΉι÷·ΦΑ»Ϊ…μΓΘ»±―ΣΓΔ‘ΌΙύΉΔΩ…÷±Ϋ”ΜρΦδΫ”ΦΛΜνKupfferœΗΑϊ(Μρ÷––‘ΑΉœΗΑϊ)Θ§Ω… ΆΖ≈Εύ÷÷œΗΑϊ“ρΉ”ΚΆΫι÷ [16]Θ§Τδ÷–Αϋά®O2¥ζ–Μ÷–Φδ≤ζΈοH2OΓΔO?2ΓΔOH?Ή‘”…ΜυΓΔO2ΦΑTNFΒ»ΓΘΔΎ”κ»±―ΣΚσΓΑΈόΙύΉΔΓ±(no-reflow)ΜρΓΑΈό―Σ‘ΌΙύΉΔΓ±(reflow-paradox)“ΐΤπΈΔ―≠ΜΖΈ…¬“”–ΙΊ [17]ΓΘΗΈώΦ «ΧΊΜ·ΒΡΟΪœΗ―ΣΙήΘ§Τδ÷±ΨΕ”κΤδΥϋΟΪœΗ―ΣΙήœύΥΤΘ§ΒΪΟΩΗωΗΈœΗΑϊΫτΝΎΝΫ…œΗΈώΦΘ§“ρΕχΕ‘ΟΪœΗ―ΣΙήΉη»ϊΒΡΡΆ ήΫœ«ΩΘ§‘Ύ…ζάμ«ιΩωœ¬“≤≥ΘΖΔ…ζΒΞΗω―ΣώΦΒΡΉη»ϊΓΘΒΪΗΈ‘ύ≥Λ ±Φδ»±―ΣΚσ”…”ΎΡΎΤΛΥ°÷ΉΓΔ―ΣώΦΡΎ―Σ“Κ≈®ΥθΦΑΈΔΨ≤¬ωΉη»ϊΒ»Θ§ Ι‘ΌΙύΉΔΚσ»±―Σ»±―θ»‘‘ΎΦΧ–χ(no-reflow)Θ§ΒΦ÷¬‘ΌΙύΉΔ≤Μ‘»Θ§Εχ‘ΌΙύΉΔΚσΓΑΈό―Σ‘ΌΙύΉΔΓ±(reflow-paradox) «”…”ΎΗ¥ΝςΜρΗ¥―θΦΛΜνΝΥΑΉœΗΑϊΘ§Ψ≠œΗΑϊ±μΟφ’≥ΗΫΖ÷Ή”ICAM-1ΓΔCD11/CD18ΓΔLΓΔP-―Γ‘ώΥΊ”κΡΎΤΛœΗΑϊάΈΙΧ’≥ΗΫΕχ“ΐΤπΗΈώΦΉη»ϊΓΘΆ®Ιΐ“‘…œΝΫ÷÷ΖΫ ΫΘ§ ΙΗ¥ΝςΚσΜν–‘―θ ΆΖ≈‘ωΕύ≤Δœό÷Τ‘ΎΨ÷≤ΩΘ§Ά§ ±Θ§”…”Ύ―θΉ‘”…ΜυΤΫΨυ ΌΟϋΚήΕΧΘ§Ϋω10?3Οκ[18]Θ§‘ΎΈΔ―≠ΜΖΈ…¬“ΒΡ«ιΩωœ¬≤ΜΡήΝΔΦ¥‘ΎΗΈΉι÷·≤Ξ…ΔΜρ¥σΝΩ ΆΖ≈»κ―Σ―≠÷–Θ§‘λ≥…Ψ÷≤ΩΉι÷·―œ÷ΊΥπ…Υ(»γΟ≈Ιή«χ)Θ§Rymsa―–ΨΩΖΔœ÷Θ§RP≥θΤΎ‘ΎΟ≈¬ω÷ήΈß«χ”ρΖ÷άκ≥ωΨΏ”–Κή«ΩΆΧ …Μν–‘ΒΡ¥σKCΘ§ΤδΉ‘ΖΔ≤ζ…ζΒΡO?2‘ωΦ”3±Ε“‘…œΓΘSuzukiΒ»‘ΎΗΈ‘ύ»±―Σ90 min‘ΌΙύ60 minΒΡΉι÷·―ßΙέ≤λ÷–ΖΔœ÷Θ§≤Γ±δ÷ς“ΣΖ÷≤Φ‘ΎΗΈ≈ίΔώ¥χ÷ΝΔρ¥χΘ§Ω…ΦϊΗΈœΗΑϊ―œ÷Ί±δ–‘Ή«÷ΉΓΔΗΈώΦπω―Σ[9]ΓΘΙΊ”ΎΤδΥπ…ΥΖ÷≤Φ≤ΩΈΜΒΡ»Ζ«–Μζ÷ΤΜΙ”–¥ΐΫχ“Μ≤Ϋ―–ΨΩΓΘ
, ΑΌΡ¥“Ϋ“©
ΒΪGonzalez-Flecha[15]―–ΨΩΖΔœ÷Θ§ΗΈœΌ≈ίΔρΓΔΔσ¥χΕ‘»±―Σ‘ΌΙύΉΔΥπ…Υ±»ΫœΟτΗ–Θ§ΡΩ«Α―–ΨΩ»œΈΣΗΈ≈ίΈό¬έ‘ΎœΗΑϊΤςΫαΙΙΜρΟΗΒΡΉιΖίΕΦ”–Υυ≤ΜΆ§Θ§’β «ΥϋΕ‘»±―Σ‘ΌΙύΉΔΥπ…ΥΟτΗ––‘≤ΜΆ§ΒΡΜυ¥ΓΓΘΡΩ«ΑΕ‘ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…ΥΟτΗ––‘ΒΡΉι÷·―ßΕ®ΈΜ»‘”–’υ“ιΘ§ΜΙ”–¥ΐΫχ“Μ≤ΫΧΫΧ÷ΓΘ
3 »±―Σ‘ΌΙύΉΔœΏΝΘΧεΚτΈϋΜν–‘ΒΡ±δΜ·
«ΑΟφ“―Ψ≠ΧαΒΫœΏΝΘΧε‘ΎœΗΑϊΗΤ≥§‘ΊΦΑ―θΉ‘”…Μυ–Έ≥…÷–ΒΡΒςΫΎΉς”ΟΓΘΒΪΒ±’β÷÷ΒςΫΎΉς”Ο≥§Ιΐ“ΜΕ®œόΕ»Θ§±Ί»Μ“ΐΤπœΏΝΘΧεΉ‘…μΥπΚΠΘ§’β÷÷ΥπΚΠ≥ωœ÷«Α Ήœ»≥ωœ÷“ΜœΒΝ–¥ζ≥Ξ–‘ΗΡ±δΘ§Ι ΝΥΫβΤδ±δΜ·Ιφ¬…Ε‘»±―Σ‘ΌΙύΉΔΥπ…ΥΒΡΖά÷ΈΨΏ”–ΜΐΦΪΒΡΉς”ΟΓΘ
ΤάΦέœΏΝΘΧεΚτΈϋΜν–‘≥Θ”ΟΚτΈϋΩΊ÷Τ¬ (RCR)ΚΆΝΉ―θ±»(P/O)ά¥±μ ΨΓΘΥϋΟ« «Ζ¥”≥œΏΝΘΧεΆξ’ϊ–‘ΓΔ―θΜ·ΝΉΥαΜ·≈ΦΝΣ≥ΧΕ»ΦΑ―θΜ·ΝΉΥαΜ·–߬ ΒΡΝιΟτ÷Η±ξΓΘΗΈ‘ύ»±―Σ‘ΌΙύΉΔ ±œΏΝΘΧεΆ®ΆΗ–‘ΒΡΗΡ±δΓΔœΏΝΘΧεΡΎΟΗΚΆΗ®ΟΗΒΡΕΣ ßΘ§“‘ΦΑ”…¥Υ‘λ≥…ΒΡ―θΜ·ΝΉΥαΜ·’œΑ≠Θ§ΨυΩ…”ΑœλœΏΝΘΧεΚτΈϋΜν–‘ΓΘIsozaki‘Ύ―–ΨΩΦδΕœΦΑ≥÷–χΗΈ‘ύ»±―ΣΕ‘œΏΝΘΧεΙΠΡήΒΡ”ΑœλΚσΖΔœ÷Θ§ΗΈ‘ύ‘Ύ≥÷–χ»±―Σ30 minΚσΕ‘RCRΦΑP/O”Αœλ≤Μ¥σΘΜ»τΫΪ»±―ΣΉή ±Φδ…ηΕ®‘Ύ60 minΘ§Ψ≠Ιέ≤λΖΔœ÷ΦδΕœ»±―Σ’ΏœΏΝΘΧεΙΠΡήΟςœ‘ΚΟ”Ύ≥÷–χ»±―Σ’ΏΘ§±μΟςΥφ≥÷–χ»±―Σ ±ΦδΒΡ―”≥ΛΘ§œΏΝΘΧεΒΡΆξ’ϊ–‘ ήΒΫ“ΜΕ®ΒΡ”ΑœλΓΘ
, ΑΌΡ¥“Ϋ“©
KorokawaΒ»―–ΨΩΗΈ¥σ≤ΩΖ÷«–≥ΐΚσ≤–ΗΈœΗΑϊœΏΝΘΧεΒΡΙΠΡή ±ΦϊΒΫΘ§ ÷ θΚσΒΎ“ΜΧλCRCΦΑR3Ϋœ ÷ θ«ΑΟςœ‘‘ωΗΏΘ§2ΓΪ5 dΚσ÷πΫΞΫΒΒΆ÷Ν θ«ΑΥ°ΤΫΘ§ΥΒΟς≤–ΗΈ¥ζ–ΜΜν‘ΨœΏΝΘΧεΙΠΡή¥ζ≥Ξ–‘‘ω«ΩΓΘΒΪKobayashi[19]Ε‘»±―Σ‘ΌΙύΉΔ ±ΗΈ‘ύœΏΝΘΧεΚτΈϋΜν–‘Ιέ≤λΖΔœ÷Θ§»±―Σ120 minœΏΝΘΧεRCRΓΔR3Ϋœ»±―Σ«ΑΟςœ‘ΫΒΒΆΘ§‘ΌΙύΉΔΚσΥδ”–ΥυΧαΗΏΘ§ΒΪ»‘Οςœ‘ΒΆ”Ύ θ«ΑΥ°ΤΫΓΘGonzalez-Flecha[10]ΒΡ―–ΨΩ”κ“‘…œœύΥΤΘ§Ιέ≤λΝΥ»±―Σ60 minΓΔ120 minΦΑ180 minΦΑ‘ΌΙύΉΔ30 min ±œΏΝΘΧεΚτΈϋΙΠΡήΒΡΗΡ±δΘ§Φϊ»±―ΣΚσRCRΓΔR3Ψυ≤ΜΆ§≥ΧΕ»ΫΒΒΆΘ§≤ΔΥφ»±―Σ ±Φδ―”≥ΛΘ§’β÷÷±δΜ·‘ΫΟςœ‘Θ§R4‘ρΈόΟςœ‘±δΜ·ΘΜ‘ΌΙύΉΔ30 min ±Θ§“‘…œ÷Η±ξ≤ΜΒΪΟΜ”–…ΐΗΏΖ¥ΕχΫœ»±―ΣΚσΟςœ‘ΫΒΒΆΘ§ΒΪ»±―Σ180 min‘ΌΙύΉΔ30 minΉι‘ρ±δΜ·≤ΜΟςœ‘ΜρΈό±δΜ·Θ§“≤–μ¥Υ ±œΏΝΘΧε¥ζ≥Ξ“―ΒΫΦΪœόΘ§Ε‘Ης÷÷¥ΧΦΛΟτΗ––‘ΫΒΒΆΥυ÷¬ΓΘ
Ής’ΏΦρΫιΘΚΚΈ–ßΕΪ(1962-)Θ§Ρ–Θ§ΥΕ ΩΘ§÷ς÷Έ“Ϋ Π
, ΑΌΡ¥“Ϋ“©
≤ΈΩΦΈΡœΉΘΚ
[1]liu W,Schob O, Pugmire JE,et al.Glycohydrolases as markers of hepatic ischemia-reperfusion injury and recovery [J].Hepatology,1996,24:157.
[2]Hatanaka N,Kamijke W,Shimizu S,et al.Ca2ΓFrelease from mitochondrial induces cytosiolic enzyme leakage in anoxis liver [J].J Surg Res,1995,58:485.
[3]Masini A,Gallesi D,Giovannini F,et al. Membrane potential of hepatic mitochondrial after cute cocaine administration in rat-the role of mitochondrial reduced glutathione [J].Hepatology,1997,25:305.
, http://www.100md.com
[4]Saris N-E L,Eriksson KD.Mitochondrial dysfunction in ischemia-reperfusion [J].Acta Anaesthesiologica Scandinavia,1995,29:(suppl 107):171.
[5]Geexaerts MD,Ronveavx-Dupal MF,Lemasters JJ,et al.Cytosilic free Ca2+and proteolysis in lethal oxidative injury in endothelial cell[J].Am J Physiol,1991,261(5):C889.
[6]Szabo I, Zoratti M.The giant channel of the inner mitochondrial membrane is inhibited by cyclosporin A [J].J Biol,Chem,1991,266:3376.
, ΑΌΡ¥“Ϋ“©
[7]Bernard P,Vassanell S,Veronese P,et al.Modulation of the mitochondrial permeability transition pore[J].J Biol Chem,1992,267:2934.
[8]Jacobs WR,Sgambati M,Gomez G,et al.Role of cytosolic Ca 2+ in renal tubule damage induced by anoxia [J].Am J Physiol,1991,260:C545.
[9]Suzuki S,Toledo-pereyra LH,Rodriguez FL,et al. Neutrophil infilration as an important factor in liver ischemia and reperfusion injury[J].Transplantation,1993,55(6):1265.
, ΑΌΡ¥“Ϋ“©
[10]Gonzalez-Flecha B, Cutrin JC,Boveris A.Time course and mechanism of oxidative stress and tissue damage in rat liver subjected to in vivo ischemia-reperfusion[J]. J Clin Invest 1993,91:456.
[11]Jaeschke H,Farbood A,Smith W.Neutrophils contribute to ischemia/reperfusion injury in rat liver in vivo [J].FASEB J ,1990,4:3355.
[12]Brass CA,Roberts TG.Hepatic free radical production after cold storage;kupffer cell-dependent and independent mechanisms in rats [J].Gastroenterology,1995,108:1167.
, http://www.100md.com
[13]Niwano M,Arii S,Monden K,et al. Aneliovation of sinusoided endothelial cell damage by kupffer cell blockade during cell preservation of rat liver [J].J Surg Res,1997,72:36.
[14]Marubayashi S,Oshire Y,Maeda T,et al.Protective effect of monoclonal antibodies to adhesion molecules on rat liver ischemia-reperfusion injury [J].Surgery,1997,122:45.
[15]Jennische E. Possible influence of glutathine on postischemic liver injury [J].Acta Pathol Microbiol Scand,[C]1984,92:55.
, ΑΌΡ¥“Ϋ“©
[16]Shibuyo H,Ohkohchi N,Seya K,et al.Kupffer cell generate superoxide anions and modulate reperfusion injury in rat liver after cold preservation [J].Hepatology,1997,25:356.
[17]Menger MD.Microcirculatory disturbance secondary to ischemia-reperfusion [J].Transplantation,1995,27(5):2863.
[18]¬§γ≤ΤΦΘ§ Ζ≥¬»ΟΘ§Άτ ΚλΘ§Β».¥σ σΗΈ»±―Σ‘ΌΙύΝςΤΎΦδ÷§÷ Ιΐ―θΜ·Ή‘”…ΜυΕ·Χ§±δΜ·ΦΑ≤ΓάμΗΡ±δΆ§≤ΫΙέ≤λ.÷–ΜΣ Β―ιΆβΩΤ‘”÷ΨΘ§1992Θ§9ΘΚ11.
[19]Kobayashi H,Nonami T,Kurokawa T,et al.Mechanism and prevention of ischemia-reperfusion liver injury in rats [J] J Surg Res,1991,51:240.
, http://www.100md.com
’Ηε»’ΤΎΘΚ1999-01-20
œΏΝΘΧε‘ΎΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡΉς”Ο
ΚΈ–ßΕΪ Ήέ ω Ε≠Φ“Κη ≤ΧΨΑ–ό …σ–Θ
’Σ“ΣΘΚΗΈ‘ύΆβΩΤΓΔ–ίΩΥΒ»≤ΓάμΙΐ≥Χ÷–ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ≤ΜΩ…±ήΟβΘ§ΗΟΙΐ≥Χ“‘»±―ΣΓΔ»±―θΓΔΡήΝΩΙ©”Π÷–ΕœΈΣ ΦΕ·“ρΥΊΘ§Ψ≠œΗΑϊΗΤ≥§‘ΊΓΔ―θΉ‘”…Μυ–Έ≥…ΦΑΑΉœΗΑϊΫΰ»σ»ΐΗω÷Ί“ΣΜΖΫΎΘ§Ά®ΙΐœΗΑϊ“ρΉ”―Ή–‘Ϋι÷ ΒΡ ΆΖ≈Θ§≤ΜΒΪΕ‘»±―ΣΗΈ‘ύ‘λ≥…Υπ…ΥΘ§Εχ«“Ε‘»Ϊ…μΤδΥϋ‘ύΤς‘λ≥…“ΜΕ®ΒΡ…ΥΚΠΘΜ‘Ύ’β“Μ≤ΓάμΙΐ≥Χ÷–Θ§œΏΝΘΧεΕ‘œΗΑϊΡΎΗΤΒΡΒςΫΎΓΔ―θΉ‘”…Μυ–Έ≥…Β»ΨΏ”–Ζ«≥Θ÷Ί“ΣΒΡΉς”ΟΓΘ»γΚΈΖά÷ΈΗΈ‘ύ»±―Σ‘ΌΙύΉΔ ±œΏΝΘΧεΥπ…ΥΫΪ «ΫώΚσ Β―ι―–ΨΩΒΡ÷ΊΒψΓΘ
ΙΊΦϋ¥ ΘΚΗΈ‘ύΦ≤≤ΓΘΜœΏΝΘΧεΘΜ »±―Σ-‘ΌΙύΉΔΥπ…Υ
÷–ΆΦΖ÷άύΚ≈ΘΚR 575 ΈΡœΉ±ξ Ε¬κΘΚA ΈΡ’¬±ύΚ≈ΘΚ1007-8622(1999)04-0295-03
, http://www.100md.com
ΗΈ‘ύ «ΧεΡΎΉν¥σΒΡ¥ζ–ΜΤςΙΌΘ§œΏΝΘΧε «œΗΑϊ¥ζ–ΜΒΡ÷––Ρ≥ΓΥυΘ§œΏΝΘΧεΒΡ’ΐ≥ΘΚτΈϋΙΠΡήΚΆATP…ζ≥…Ε‘Ήι÷·ΤςΙΌΙΠΡήΦΑœΗΑϊΫαΙΙΒΡΆξ’ϊ–‘÷ΝΙΊ÷Ί“ΣΓΘΒΪœΏΝΘΧεΕ‘»±―ΣΓΔ»±―θΖ«≥ΘΟτΗ–Θ§»±―ΣΓΔ»±―θΒΡΗΈΉι÷·‘Ύ―ΣΝςΜ÷Η¥ΜρΗ¥―θΚσΤδΥπ…Υ≥ΧΕ»≤ΜΒΪΟΜ”–ΗΡ…ΤΘ§Ζ¥ΕχΫχ“Μ≤ΫΦ”÷ΊΘ§’β”κœΗΑϊΗΤ≥§‘ΊΓΔ―θΉ‘”…ΜυΒ»“ρΥΊ”–÷±Ϋ”ΒΡΙΊœΒ[1]ΓΘœΏΝΘΧε «ΝΣœΒΗΤ≥§‘ΊΓΔ―θΉ‘”…Μυ–Έ≥…ΚΆœΗΑϊΥάΆωΒΡ÷––ΡΜΖΫΎΓΘΫϋΡξΕ‘œΏΝΘΧε‘ΎΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡΉς”Ο―–ΨΩΫœΕύΘ§Ϋχ’ΙΫœΩλΘ§±ΨΈΡΨΆ’βΖΫΟφΒΡΡΎ»ίΉω“ΜΉέ ωΓΘ
1 œΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ”κ»±―Σ‘ΌΙύΉΔΗΤ≥§‘ΊΒΡΝΣœΒ
ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΩ…÷¬œΗΑϊΗΤ≥§‘ΊΘ§ «“ΐΤπΗΈœΗΑϊΥπ…ΥΒΡ‘≠“ρ÷°“ΜΘ§ΕχœΏΝΘΧεΨΏ”–«Ω¥σΒΡΨέΜΐCa2+ΡήΝΠΘ§Ε‘ΗΤάκΉ”Έ»Χ§ΤπΒΫ“ΜΕ®ΒΡΒςΫΎΉς”ΟΘ§ΒΪœΏΝΘΧε…ψCa2+ΡήΝΠ”–œόΘ§»τΥπ…Υ“ρΥΊ≤ΜΡή«ε≥ΐΘ§Ψ≠Ης÷÷ΆΨΨΕΉν÷’Ω…÷¬œΗΑϊΡΎΗΤ≥§‘Ί[2Θ§3]ΓΘ
ΫϋΡξΕ‘œΗΑϊΜΒΥάΒΡ―–ΨΩ±μΟςΘ§œΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ(Mitochondrial membrane permeability transition,MMPT)‘ΎœΗΑϊΥάΆω÷–ΤπΉ≈ΙΊΦϋΒΡΉς”Ο[4]ΓΘ‘ΎΗς÷÷Υπ…Υ“ρΥΊΒΡΉς”Οœ¬Θ§MMPT“ΜΒ©ΖΔ…ζΘ§œΏΝΘΧεΡΛΆ®ΆΗ–‘―ΗΥΌ…ΐΗΏΘ§œΗΑϊΥάΆωΥφ÷°ΖΔ…ζΓΘ“÷÷ΤMMPTΖΔ…ζΩ…―”ΜΚœΗΑϊΥάΆω[5]ΓΘΡΩ«Α»œΈΣMMPTΒΡΖΔ…ζΜζ÷ΤΑϋά®œΏΝΘΧεΡΎΡΛΒΡΖ«ΧΊ“λ–‘Υπ…ΥΚΆ“Μ÷÷ΨόΆ®Βά(Megapore)ΒΡΩΣΖ≈[4Θ§6]ΓΘΕχœΏΝΘΧεΡΎΡΛΖ«ΧΊ“λ–‘Υπ…ΥΩ…”…ΗΤ≥§‘Ί¥ΞΖΔΒΡ―θΉ‘”…ΜυΥπ…ΥΒ»‘λ≥…ΓΘΙΊ”ΎΒΦ÷¬MMPTΒΡΧΊ βΜζ÷ΤΒΡ―–ΨΩ±μΟςΘΚœΏΝΘΧεΜυ≤ψ÷–Ca2ΓFΒΡ¥σΝΩΨέΜΐ’ΐ «ΨόΆ®Βά(Megapore)ΩΣΖ≈ΒΡ‘≠“ρ÷°“Μ[5Θ§6]ΓΘΙ œΏΝΘΧεΗΤ≥§‘ΊΆ®ΙΐMMPTΩ…ΒΦ÷¬œΗΑϊ÷¬Υά–‘Υπ…ΥΓΘ
, ΑΌΡ¥“Ϋ“©
Ε‘ΨόΆ®ΒάΫχ“Μ≤Ϋ―–ΨΩΖΔœ÷Θ§œΏΝΘΧεΡΛ…œΒΡΨόΆ®ΒάΨΆ «÷±ΨΕΈΣ20
, ΑΌΡ¥“Ϋ“©
ΒΪΝμ”–―–ΨΩ»œΈΣΘ§‘ΎœΗΑϊΥπ…Υ÷–Θ§œΏΝΘΧε…ψΗΤ‘ωΕύ“≤–μ «ΦΧΖΔ–‘ΗΡ±δΒΡΫαΙϊΘ§ΕχœΏΝΘΧεΡΛΆ®ΆΗ–‘ΗΡ±δ(MMPT)≤≈ « ΦΕ·“ρΥΊΘ§“ρΕχΕ‘ΑϊΫ§ΗΤ≥§‘Ί‘ΎΉι÷·»±―Σ‘ΌΙύΉΔΥπ…Υ÷–ΒΡ÷Ί“Σ–‘Χα≥ω÷ “…Θ§[8]Θ§“ρΈΣ‘Ύ¥σ σΗΈ‘ύ»±―θ35 min‘ΌΙύΉΔ10 minΒΡΡΘ–Ά÷–Θ§ΗΈΉι÷·ΒγΨΒœ¬Ά§“Μ ”“ΑΒΡΝΫΗωΗΈœΗΑϊ÷–Θ§“ΜΗωœΏΝΘΧε’ΐ≥ΘΘ§ΕχΝμ“ΜΗω‘ρΥ°÷Ή±δ–‘Θ§Κσ’Ώ“≤–μ“―”–œΏΝΘΧεΆ®ΆΗ–‘ΗΡ±δΘΜΕχ÷ΒΒΟΉΔ“βΒΡ «¥Υ ±»‘Έ¥ΦϊΤδΥϋœΗΑϊΤς”––ΈΧ§―ßΖΫΟφΒΡ±δΜ·[11]ΓΘ’βΫω «≥θ≤ΫΙέ≤λΒΡΫαΙϊΘ§“Σ≤ϊΟςΤδœξœΗΜζ÷ΤΘ§ΜΙ”–¥ΐΗϋΫχ“Μ≤Ϋ―–ΨΩΓΘ
2 œΏΝΘΧε‘Ύ―θΉ‘”…Μυ–Έ≥…÷–ΒΡΉς”Ο
ΜΤύ―Ώ ―θΜ·ΟΗΚΆ÷––‘ΑΉœΗΑϊ «―θΉ‘”…ΜυΒΡΝΫΗω÷Ί“Σά¥‘¥Θ§[9]Θ§»±―ΣΚΆ‘ΌΙύΉΔΨυΩ…≤ζ…ζΫœΕύ≥§―θΜ·ΈοΘ§“ΐΤπΉι÷·Υπ…ΥΓΘœΏΝΘΧε «ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΤΎΦδΜν–‘―θ≤ζ…ζΒΡ÷Ί“ΣΤςΙΌ[10]ΓΘ
2.1 ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΜν–‘―θ ΆΖ≈ΧΊΒψ”κΗΈœΗΑϊΥπ…ΥΒΡΙΊœΒΘΚΕ‘ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…Υ―–ΨΩ±μΟςΘ§Η¥―θΚσΜν–‘―θ ΆΖ≈ ±œύ”κΗΈœΗΑϊΥπ…ΥΟή«–œύΙΊΓΘJaeschkeΒ»[11]ΉηΕœ≤ΩΖ÷ΗΈ‘ύ―ΣΝς45 minΚσ‘ΌΙύΉΔ(RP)≤ΜΆ§ ±ΦδΘ§ΖΔœ÷ΗΈΉι÷·≥ωœ÷ΝΥΥΪœύΥπ…ΥΓΘ‘ΎΜ÷Η¥ΙύΉΔΒΡ1 hΡΎΘ§―ΣΫ§ΉΣΑ±ΟΗΜν–‘…ΐΗΏΘ§¥ΥΈΣΗΈ‘ύ»±―Σ‘ΌΙύΉΔΒΡΔώœύΥπ…Υ(initial injury)ΘΜΫ”œ¬»ΞΒΡ5 h Θ§ΉΣΑ±ΟΗΒΡΜν–‘Έ§≥÷‘ΎΔώœύΥπ…ΥΥ°ΤΫΕχ‘ΎRPΚσ6ΓΪ24 hΘ§ΉΣΑ±ΟΗΜν–‘‘Ό¥Έœ‘÷χ…ΐΗΏΘ§±μΟς»±―Σ‘ΌΙύΉΔΗΈ‘ύΜΙ¥φ‘ΎΔρœύΥπ…Υ(progressive injury)ΓΘ¥”ΕχΩ…“‘Ω¥≥ωΗ¥―θ ±ΗΈœΗΑϊΥπ…Υ”κΜν–‘―θΒΡ ΆΖ≈ ±œύΨΏ”–÷±Ϋ”ΝΣœΒΓΘ Β―ιΜΙ÷ΛΟςRPΗΈ‘ύΒΡΔώœύΥπ…Υ”κΗΈώΦΩίΖώ œœΗΑϊ(KC)”–ΙΊΓΘKCΩ…“ρΗΤ≥§‘ΊΓΔΆΧ …Ής”ΟΜρ≤ΙΧεœΒΆ≥ΕχΦΛΜνΘ§ΜνΜ·ΒΡKC≥ΐ÷ς“Σ ΆΖ≈Μν–‘―θΆβΘ§ΜΙΩ… ΆΖ≈ΒΑΑΉΟΗΦΑΗς÷÷œΗΑϊ“ρΉ”Θ§ΫιΒΦΕ‘ΗΈœΗΑϊΒΡΕΨ–‘Ής”ΟΘ§ΒΦ÷¬―œ÷ΊΒΡΗΈ‘ύ‘ΌΙύΉΔΥπ…ΥΘΜΗ…»≈KCΙΠΡήΒΡ“©ΈοΨΏ”–Οςœ‘ΒΡ÷ΈΝΤΉς”Ο[12Θ§13]Θ§ΕχΔρœύΥπ…Υ”κΑΉœΗΑϊ‘ΎΨ÷≤ΩΒΡΨέΦ·ΒΫ“ΜΕ®ΝΩ≤ΔΦΛΜνΚσ ΆΖ≈ΕΨ–‘Ϋι÷ ”–ΙΊΘ§ΨΓΙήΑΉœΗΑϊ‘ΎRP≥θΤΎ‘ΎΗΈώΦ”κΡΎΤΛœΗΑϊ’≥ΗΫ‘ω«Ω≤ΔΩΣ ΦΨέΦ·Θ§ΒΪ≤Δ≤Μ≤Έ”κΗΈΉι÷·ΔώœύΥπ…Υ[13]ΓΘΆ§ ±ΜΙΖΔœ÷KCΒΡΔώœύΥπ…ΥΩ…ΦΛΜν÷––‘ΑΉœΗΑϊ(PMN)Θ§ ΙΤδ ΆΖ≈Μν–‘―θΦΑΒΑΑΉΟΗΒ»ΕΨ–‘Ϋι÷ ΙΞΜςΗΈœΗΑϊΘ§ΜΒΥάΗΈœΗΑϊΖ¥Ιΐά¥”÷Ω…¥ΌΫχ≤ΙΧεΦΛΜ·ΚΆPMNΜνΜ·Θ§»γ¥ΥΕώ–‘―≠ΜΖΘ§Ήν÷’ΒΦ÷¬―œ÷ΊΗΈœΗΑϊΥπ…ΥΓΘΒΞΩΥ¬ΓΩΙΧεΩ…Φθ…ΌPMN’≥ΗΫ≤Δ Ι‘ΌΙύΉΔΥπ…ΥΦθ«αΓΘ[14]ΓΘ
, http://www.100md.com
2.2 Μν–‘―θ”κ»±―Σ‘ΌΙύΉΔΚσΥπ…ΥΗΈœΗΑϊ‘ΎΗΈœΌ≈ί÷–ΒΡΖ÷≤ΦΘΚ“‘«Α―–ΨΩΖΔœ÷Θ§ΗΈ“Ε«–≥ΐΚσΗΈœΗΑϊΒΡ‘Ό…ζ÷ς“ΣΖΔ…ζ‘ΎΗΈ≈ίΔώ¥χΘ§ΕχΫα‘ζΕ·ΈοΗΈ―ΣΙή ΙΗΈ‘ύ»±―θΜρ”Σ―χ≤ΜΝΦ–‘‘γΤΎΗΈ”≤Μ·ΥυΖΔ…ζΒΡΗΈΥπΚΠ÷ς“ΣΖΔ…ζ‘ΎΔσ¥χΘ§’βΩ…Ϋβ ΆΈΣΔώ¥χΗΈœΗΑϊΩΩΫϋ÷–÷α―ΣΙήΘ§”≈œ»ΜώΒΟΗΜ”Ύ”Σ―χΒΡΈο÷ ΚΆΚ§―θ≥δΉψΒΡ―Σ“ΚΘ§“ρ¥Υ”≈œ»‘Ό…ζΘ§ΕχΔσ¥χΉι÷·PO2ΉνΒΆΘ§Ι Ήν»ί“Ή≥ωœ÷»±―θ–‘ΗΈΥπΚΠΚΆΗΈœΗΑϊΥάΆωΓΘ
Ρ«Ο¥Θ§ΗΈ‘ύ»±―ΣΚσ‘ΌΙύΉΔΥπ…ΥΒΡΧΊΒψ « ≤Ο¥ΡΊ?FlreemanΒ»ΒΡ―–ΨΩ»œΈΣΘΚΜΙ‘≠–ΆΙ»κΉΗ κΡ(GSH)‘ΎΗΈ‘ύ «÷Ί“ΣΒΡΉ‘”…Μυ«ε≥ΐΦΝΓΘ―–ΨΩΖΔœ÷Θ§¥σ σΗΈ‘ύ»±―Σ90 minΚσ‘ΌΙύΉΔΘ§ΗΈΉι÷·GSHΒΆ’Ώ(»γΫϊ ≥)ΗΈ‘ύΥπΚΠ≥ΧΕ»ΫœGSHΗΏ’ΏΗϋΈΣ―œ÷ΊΘ§≤Δ«“ΥπΚΠΗΈœΗΑϊ÷ς“ΣΖ÷≤Φ‘Ύ–Γ“ΕΦδΟ≈Ψ≤¬ω÷ήΈß«χ”ρ(Φ¥Δώ¥χ)Θ§’β÷÷±μœ÷‘ΎGSHΆξ»ΪœϊΚΡ’ΏΗϋΦ”Οςœ‘Θ§“ρΕχ÷ß≥÷―θΉ‘”…Μυ“ΐΤπΗΈΥπ…Υ’β“Μ¬έΕœ[15]ΓΘΆ§ ± Β―ι“≤÷ΛΟςΘ§”Ο¬»Μ·νή¥ΠάμΫϊ ≥ΒΡΕ·ΈοΘ§“‘‘ωΦ”GSH‘ΎΗΈΉι÷·ΒΡΚ§ΝΩΘ§ΫαΙϊ»±―ΣΚσΗΈœΗΑϊΜΒΥά≥ΧΕ»Οςœ‘Φθ«α[15]ΘΜ’β÷÷Ε‘»±―Σ‘ΌΙύΉΔΥπ…ΥΒΡ«χ¥χ–‘≤ν“λΘ§Ω…Ρή”κ“‘œ¬“ρΥΊ”–ΙΊΘΚΔΌ”κKupfferœΗΑϊ‘ΎΗΈώΦΒΡ≤ΜΨυ÷ –‘Ζ÷≤Φ”–ΙΊΓΘ¥σ σΗΈKupfferœΗΑϊ‘Φ’ΦΗΈ Β÷ ΒΡ2.1%Θ§ΒΪ“‘ΗΈ–Γ“Ε÷ή±Ώ¥χΖ÷≤ΦΫœΕύΓΘkupfferœΗΑϊΒΡΜνΜ·‘ΎΜζΧεΖάΈάΜζΡή÷–”–ΜΐΦΪΒΡΉς”ΟΘ§ΒΪΜνΜ·ΚσΒΡKupfferœΗΑϊ“‘ΤδΖ÷ΟΎΒΡΗς÷÷œΗΑϊ“ρΉ”(cytotines)Ής”Ο”ΎΗΈœΗΑϊΓΔΝΎΫϋΉι÷·ΦΑ»Ϊ…μΓΘ»±―ΣΓΔ‘ΌΙύΉΔΩ…÷±Ϋ”ΜρΦδΫ”ΦΛΜνKupfferœΗΑϊ(Μρ÷––‘ΑΉœΗΑϊ)Θ§Ω… ΆΖ≈Εύ÷÷œΗΑϊ“ρΉ”ΚΆΫι÷ [16]Θ§Τδ÷–Αϋά®O2¥ζ–Μ÷–Φδ≤ζΈοH2OΓΔO?2ΓΔOH?Ή‘”…ΜυΓΔO2ΦΑTNFΒ»ΓΘΔΎ”κ»±―ΣΚσΓΑΈόΙύΉΔΓ±(no-reflow)ΜρΓΑΈό―Σ‘ΌΙύΉΔΓ±(reflow-paradox)“ΐΤπΈΔ―≠ΜΖΈ…¬“”–ΙΊ [17]ΓΘΗΈώΦ «ΧΊΜ·ΒΡΟΪœΗ―ΣΙήΘ§Τδ÷±ΨΕ”κΤδΥϋΟΪœΗ―ΣΙήœύΥΤΘ§ΒΪΟΩΗωΗΈœΗΑϊΫτΝΎΝΫ…œΗΈώΦΘ§“ρΕχΕ‘ΟΪœΗ―ΣΙήΉη»ϊΒΡΡΆ ήΫœ«ΩΘ§‘Ύ…ζάμ«ιΩωœ¬“≤≥ΘΖΔ…ζΒΞΗω―ΣώΦΒΡΉη»ϊΓΘΒΪΗΈ‘ύ≥Λ ±Φδ»±―ΣΚσ”…”ΎΡΎΤΛΥ°÷ΉΓΔ―ΣώΦΡΎ―Σ“Κ≈®ΥθΦΑΈΔΨ≤¬ωΉη»ϊΒ»Θ§ Ι‘ΌΙύΉΔΚσ»±―Σ»±―θ»‘‘ΎΦΧ–χ(no-reflow)Θ§ΒΦ÷¬‘ΌΙύΉΔ≤Μ‘»Θ§Εχ‘ΌΙύΉΔΚσΓΑΈό―Σ‘ΌΙύΉΔΓ±(reflow-paradox) «”…”ΎΗ¥ΝςΜρΗ¥―θΦΛΜνΝΥΑΉœΗΑϊΘ§Ψ≠œΗΑϊ±μΟφ’≥ΗΫΖ÷Ή”ICAM-1ΓΔCD11/CD18ΓΔLΓΔP-―Γ‘ώΥΊ”κΡΎΤΛœΗΑϊάΈΙΧ’≥ΗΫΕχ“ΐΤπΗΈώΦΉη»ϊΓΘΆ®Ιΐ“‘…œΝΫ÷÷ΖΫ ΫΘ§ ΙΗ¥ΝςΚσΜν–‘―θ ΆΖ≈‘ωΕύ≤Δœό÷Τ‘ΎΨ÷≤ΩΘ§Ά§ ±Θ§”…”Ύ―θΉ‘”…ΜυΤΫΨυ ΌΟϋΚήΕΧΘ§Ϋω10?3Οκ[18]Θ§‘ΎΈΔ―≠ΜΖΈ…¬“ΒΡ«ιΩωœ¬≤ΜΡήΝΔΦ¥‘ΎΗΈΉι÷·≤Ξ…ΔΜρ¥σΝΩ ΆΖ≈»κ―Σ―≠÷–Θ§‘λ≥…Ψ÷≤ΩΉι÷·―œ÷ΊΥπ…Υ(»γΟ≈Ιή«χ)Θ§Rymsa―–ΨΩΖΔœ÷Θ§RP≥θΤΎ‘ΎΟ≈¬ω÷ήΈß«χ”ρΖ÷άκ≥ωΨΏ”–Κή«ΩΆΧ …Μν–‘ΒΡ¥σKCΘ§ΤδΉ‘ΖΔ≤ζ…ζΒΡO?2‘ωΦ”3±Ε“‘…œΓΘSuzukiΒ»‘ΎΗΈ‘ύ»±―Σ90 min‘ΌΙύ60 minΒΡΉι÷·―ßΙέ≤λ÷–ΖΔœ÷Θ§≤Γ±δ÷ς“ΣΖ÷≤Φ‘ΎΗΈ≈ίΔώ¥χ÷ΝΔρ¥χΘ§Ω…ΦϊΗΈœΗΑϊ―œ÷Ί±δ–‘Ή«÷ΉΓΔΗΈώΦπω―Σ[9]ΓΘΙΊ”ΎΤδΥπ…ΥΖ÷≤Φ≤ΩΈΜΒΡ»Ζ«–Μζ÷ΤΜΙ”–¥ΐΫχ“Μ≤Ϋ―–ΨΩΓΘ
, ΑΌΡ¥“Ϋ“©
ΒΪGonzalez-Flecha[15]―–ΨΩΖΔœ÷Θ§ΗΈœΌ≈ίΔρΓΔΔσ¥χΕ‘»±―Σ‘ΌΙύΉΔΥπ…Υ±»ΫœΟτΗ–Θ§ΡΩ«Α―–ΨΩ»œΈΣΗΈ≈ίΈό¬έ‘ΎœΗΑϊΤςΫαΙΙΜρΟΗΒΡΉιΖίΕΦ”–Υυ≤ΜΆ§Θ§’β «ΥϋΕ‘»±―Σ‘ΌΙύΉΔΥπ…ΥΟτΗ––‘≤ΜΆ§ΒΡΜυ¥ΓΓΘΡΩ«ΑΕ‘ΗΈ‘ύ»±―Σ‘ΌΙύΉΔΥπ…ΥΟτΗ––‘ΒΡΉι÷·―ßΕ®ΈΜ»‘”–’υ“ιΘ§ΜΙ”–¥ΐΫχ“Μ≤ΫΧΫΧ÷ΓΘ
3 »±―Σ‘ΌΙύΉΔœΏΝΘΧεΚτΈϋΜν–‘ΒΡ±δΜ·
«ΑΟφ“―Ψ≠ΧαΒΫœΏΝΘΧε‘ΎœΗΑϊΗΤ≥§‘ΊΦΑ―θΉ‘”…Μυ–Έ≥…÷–ΒΡΒςΫΎΉς”ΟΓΘΒΪΒ±’β÷÷ΒςΫΎΉς”Ο≥§Ιΐ“ΜΕ®œόΕ»Θ§±Ί»Μ“ΐΤπœΏΝΘΧεΉ‘…μΥπΚΠΘ§’β÷÷ΥπΚΠ≥ωœ÷«Α Ήœ»≥ωœ÷“ΜœΒΝ–¥ζ≥Ξ–‘ΗΡ±δΘ§Ι ΝΥΫβΤδ±δΜ·Ιφ¬…Ε‘»±―Σ‘ΌΙύΉΔΥπ…ΥΒΡΖά÷ΈΨΏ”–ΜΐΦΪΒΡΉς”ΟΓΘ
ΤάΦέœΏΝΘΧεΚτΈϋΜν–‘≥Θ”ΟΚτΈϋΩΊ÷Τ¬ (RCR)ΚΆΝΉ―θ±»(P/O)ά¥±μ ΨΓΘΥϋΟ« «Ζ¥”≥œΏΝΘΧεΆξ’ϊ–‘ΓΔ―θΜ·ΝΉΥαΜ·≈ΦΝΣ≥ΧΕ»ΦΑ―θΜ·ΝΉΥαΜ·–߬ ΒΡΝιΟτ÷Η±ξΓΘΗΈ‘ύ»±―Σ‘ΌΙύΉΔ ±œΏΝΘΧεΆ®ΆΗ–‘ΒΡΗΡ±δΓΔœΏΝΘΧεΡΎΟΗΚΆΗ®ΟΗΒΡΕΣ ßΘ§“‘ΦΑ”…¥Υ‘λ≥…ΒΡ―θΜ·ΝΉΥαΜ·’œΑ≠Θ§ΨυΩ…”ΑœλœΏΝΘΧεΚτΈϋΜν–‘ΓΘIsozaki‘Ύ―–ΨΩΦδΕœΦΑ≥÷–χΗΈ‘ύ»±―ΣΕ‘œΏΝΘΧεΙΠΡήΒΡ”ΑœλΚσΖΔœ÷Θ§ΗΈ‘ύ‘Ύ≥÷–χ»±―Σ30 minΚσΕ‘RCRΦΑP/O”Αœλ≤Μ¥σΘΜ»τΫΪ»±―ΣΉή ±Φδ…ηΕ®‘Ύ60 minΘ§Ψ≠Ιέ≤λΖΔœ÷ΦδΕœ»±―Σ’ΏœΏΝΘΧεΙΠΡήΟςœ‘ΚΟ”Ύ≥÷–χ»±―Σ’ΏΘ§±μΟςΥφ≥÷–χ»±―Σ ±ΦδΒΡ―”≥ΛΘ§œΏΝΘΧεΒΡΆξ’ϊ–‘ ήΒΫ“ΜΕ®ΒΡ”ΑœλΓΘ
, ΑΌΡ¥“Ϋ“©
KorokawaΒ»―–ΨΩΗΈ¥σ≤ΩΖ÷«–≥ΐΚσ≤–ΗΈœΗΑϊœΏΝΘΧεΒΡΙΠΡή ±ΦϊΒΫΘ§ ÷ θΚσΒΎ“ΜΧλCRCΦΑR3Ϋœ ÷ θ«ΑΟςœ‘‘ωΗΏΘ§2ΓΪ5 dΚσ÷πΫΞΫΒΒΆ÷Ν θ«ΑΥ°ΤΫΘ§ΥΒΟς≤–ΗΈ¥ζ–ΜΜν‘ΨœΏΝΘΧεΙΠΡή¥ζ≥Ξ–‘‘ω«ΩΓΘΒΪKobayashi[19]Ε‘»±―Σ‘ΌΙύΉΔ ±ΗΈ‘ύœΏΝΘΧεΚτΈϋΜν–‘Ιέ≤λΖΔœ÷Θ§»±―Σ120 minœΏΝΘΧεRCRΓΔR3Ϋœ»±―Σ«ΑΟςœ‘ΫΒΒΆΘ§‘ΌΙύΉΔΚσΥδ”–ΥυΧαΗΏΘ§ΒΪ»‘Οςœ‘ΒΆ”Ύ θ«ΑΥ°ΤΫΓΘGonzalez-Flecha[10]ΒΡ―–ΨΩ”κ“‘…œœύΥΤΘ§Ιέ≤λΝΥ»±―Σ60 minΓΔ120 minΦΑ180 minΦΑ‘ΌΙύΉΔ30 min ±œΏΝΘΧεΚτΈϋΙΠΡήΒΡΗΡ±δΘ§Φϊ»±―ΣΚσRCRΓΔR3Ψυ≤ΜΆ§≥ΧΕ»ΫΒΒΆΘ§≤ΔΥφ»±―Σ ±Φδ―”≥ΛΘ§’β÷÷±δΜ·‘ΫΟςœ‘Θ§R4‘ρΈόΟςœ‘±δΜ·ΘΜ‘ΌΙύΉΔ30 min ±Θ§“‘…œ÷Η±ξ≤ΜΒΪΟΜ”–…ΐΗΏΖ¥ΕχΫœ»±―ΣΚσΟςœ‘ΫΒΒΆΘ§ΒΪ»±―Σ180 min‘ΌΙύΉΔ30 minΉι‘ρ±δΜ·≤ΜΟςœ‘ΜρΈό±δΜ·Θ§“≤–μ¥Υ ±œΏΝΘΧε¥ζ≥Ξ“―ΒΫΦΪœόΘ§Ε‘Ης÷÷¥ΧΦΛΟτΗ––‘ΫΒΒΆΥυ÷¬ΓΘ
Ής’ΏΦρΫιΘΚΚΈ–ßΕΪ(1962-)Θ§Ρ–Θ§ΥΕ ΩΘ§÷ς÷Έ“Ϋ Π
, http://www.100md.com
Ής’ΏΒΞΈΜΘΚΚΈ–ßΕΪΘΚΫβΖ≈ΨϋΒΎ“Μ“Ϋ‘ΚΗΈΒ®ΆβΩΤ÷––ΡΘ§άΦ÷ί730030
Ε≠Φ“Κη ≤ΧΨΑ–όΘΚΒΎ»ΐΨϋ“Ϋ¥σ―ßΈςΡœ“Ϋ‘ΚΗΈΒ®ΆβΩΤ÷––ΡΘ§÷Ί«λ 400038
≤ΈΩΦΈΡœΉΘΚ
[1]liu W,Schob O, Pugmire JE,et al.Glycohydrolases as markers of hepatic ischemia-reperfusion injury and recovery [J].Hepatology,1996,24:157.
[2]Hatanaka N,Kamijke W,Shimizu S,et al.Ca2ΓFrelease from mitochondrial induces cytosiolic enzyme leakage in anoxis liver [J].J Surg Res,1995,58:485.
, ΑΌΡ¥“Ϋ“©
[3]Masini A,Gallesi D,Giovannini F,et al. Membrane potential of hepatic mitochondrial after cute cocaine administration in rat-the role of mitochondrial reduced glutathione [J].Hepatology,1997,25:305.
[4]Saris N-E L,Eriksson KD.Mitochondrial dysfunction in ischemia-reperfusion [J].Acta Anaesthesiologica Scandinavia,1995,29:(suppl 107):171.
[5]Geexaerts MD,Ronveavx-Dupal MF,Lemasters JJ,et al.Cytosilic free Ca2+and proteolysis in lethal oxidative injury in endothelial cell[J].Am J Physiol,1991,261(5):C889.
, http://www.100md.com
[6]Szabo I, Zoratti M.The giant channel of the inner mitochondrial membrane is inhibited by cyclosporin A [J].J Biol,Chem,1991,266:3376.
[7]Bernard P,Vassanell S,Veronese P,et al.Modulation of the mitochondrial permeability transition pore[J].J Biol Chem,1992,267:2934.
[8]Jacobs WR,Sgambati M,Gomez G,et al.Role of cytosolic Ca 2+ in renal tubule damage induced by anoxia [J].Am J Physiol,1991,260:C545.
, ΑΌΡ¥“Ϋ“©
[9]Suzuki S,Toledo-pereyra LH,Rodriguez FL,et al. Neutrophil infilration as an important factor in liver ischemia and reperfusion injury[J].Transplantation,1993,55(6):1265.
[10]Gonzalez-Flecha B, Cutrin JC,Boveris A.Time course and mechanism of oxidative stress and tissue damage in rat liver subjected to in vivo ischemia-reperfusion[J]. J Clin Invest 1993,91:456.
[11]Jaeschke H,Farbood A,Smith W.Neutrophils contribute to ischemia/reperfusion injury in rat liver in vivo [J].FASEB J ,1990,4:3355.
, http://www.100md.com
[12]Brass CA,Roberts TG.Hepatic free radical production after cold storage;kupffer cell-dependent and independent mechanisms in rats [J].Gastroenterology,1995,108:1167.
[13]Niwano M,Arii S,Monden K,et al. Aneliovation of sinusoided endothelial cell damage by kupffer cell blockade during cell preservation of rat liver [J].J Surg Res,1997,72:36.
[14]Marubayashi S,Oshire Y,Maeda T,et al.Protective effect of monoclonal antibodies to adhesion molecules on rat liver ischemia-reperfusion injury [J].Surgery,1997,122:45.
, ΑΌΡ¥“Ϋ“©
[15]Jennische E. Possible influence of glutathine on postischemic liver injury [J].Acta Pathol Microbiol Scand,[C]1984,92:55.
[16]Shibuyo H,Ohkohchi N,Seya K,et al.Kupffer cell generate superoxide anions and modulate reperfusion injury in rat liver after cold preservation [J].Hepatology,1997,25:356.
[17]Menger MD.Microcirculatory disturbance secondary to ischemia-reperfusion [J].Transplantation,1995,27(5):2863.
[18]¬§γ≤ΤΦΘ§ Ζ≥¬»ΟΘ§Άτ ΚλΘ§Β».¥σ σΗΈ»±―Σ‘ΌΙύΝςΤΎΦδ÷§÷ Ιΐ―θΜ·Ή‘”…ΜυΕ·Χ§±δΜ·ΦΑ≤ΓάμΗΡ±δΆ§≤ΫΙέ≤λ.÷–ΜΣ Β―ιΆβΩΤ‘”÷ΨΘ§1992Θ§9ΘΚ11.
[19]Kobayashi H,Nonami T,Kurokawa T,et al.Mechanism and prevention of ischemia-reperfusion liver injury in rats [J] J Surg Res,1991,51:240.
’Ηε»’ΤΎΘΚ1999-01-20, ΑΌΡ¥“Ϋ“©