表达FasL的肝癌细胞诱导淋巴细胞凋亡肿瘤细胞的反击免疫机制
作者:王晋豫 骆抗先 陈泊 何海棠 朱幼芙 胡琼
单位:510515 广州,第一军医大学南方医院
关键词:
中华医学杂志980807 我们应用免疫组织化学法发现肝癌病人的癌细胞Fas配体(FasL)阳性而汇管区浸润淋巴细胞发生凋亡[1]。为探讨FasL在肿瘤免疫逃避中的作用,用抗癌药诱导肝癌细胞株HepG2表达FasL,再与Jurkat T淋巴细胞共培养,经流式细胞仪研究其相互作用。
一、材料和方法
1.药物诱导实验:HepG2是1979年建立的人肝癌细胞株,Jurkat为人T淋巴细胞白血病细胞株,高水平表达Fas并对FasL及抗Fas单克隆抗体诱导的凋亡敏感(中国协和医科大学肿瘤研究所张叔人教授惠赠)。于HepG2细胞株中分别加入0、10、50、100、200 μg/ml甲氨喋呤12~24小时,收集细胞及上清液。常规提取RNA,用反义链引物合成cDNA。引物FasL-5′ATGTTTCAGCTCTTCCACCTACA GAAGGA3′和FasLR5′-TGACCAGAGAGAGCTCAGATACGTTGAC3′;β-actin-5′TGACGGGGTCACCCACACTGTGCCCATCT A3′和β-actinR-5′TCCTTAATGTCA CGCTCACGATTTC3′。常规PCR步骤。阳性对照用FasL质粒(广州中山医科大学徐安龙教授惠赠)。
, 百拇医药
2.共培养试验:HepG2细胞贴壁后加甲氨喋呤,孵育24小时去除上清液,PBS洗3次;5×106/ml Jurkat加入10 μmol/L 5-溴-2-脱氧尿嘧啶(BrdU)预孵育24小时作内源性DNA标志。每瓶(25 cm2)HepG2加入2.5×106 Jurkat,孵育24小时。HepG2加2 μg/ml抗FasL单克隆抗体,37 ℃温育1小时再加入Jurkat,作为FasL/Fas介导凋亡的特异阻断试验。HepG2处理后洗3次再孵育24小时,收集上清加入Jurkat培养48小时,观察上清中FasL活性。5×106/ml Jurkat细胞加入100 ng/ml抗Fas单克隆抗体(IgG3),为Fas介导凋亡的对照实验。
3.凋亡细胞的流式细胞仪检测:收集培养细胞,加入鼠抗BrdU抗体50 μl,再加抗鼠IgG异硫氰基荧光素(FITC)50 μl, 37℃各孵育30分钟;然后加含1%RNA 酶的Tris-HCl缓冲液500 μl孵育30分钟,加碘化丙啶(PI)进行DNA染色上机分析。
, 百拇医药
二、结果
1.药物诱导HepG2表达FasL:见图1。
注:M:DNA分子量标志;1.FasL质粒PCR产物;2.未诱导的HepG2细胞;3~6:HepG2细胞
图1 HepG2细胞经甲氨喋呤诱导表达
FasL mRNA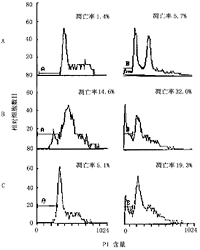
A.未诱导的HepG2与Jurkat共培养,图中A、B为本底凋亡值;B.诱导后的HepG2杀伤Jurkat细胞:出现凋亡的亚G1峰,HepG2被抗癌药杀伤,Jurkat被HepG2杀伤;C.HepG2经氨甲喋呤诱导,抗FasL阻断,再植入Jurkat, Jurkat仅有本底凋亡,HepG2凋亡率下降图2 氨甲喋呤诱导表达FasL的HepG2细胞对Jurkat细胞的杀伤
, 百拇医药
2.共培养及凋亡检测:以BrdU预标记Jurkat(标记率95%)、将2种细胞标记FITC,除去FITC低值的HepG2,分析FITC高值Jurkat的细胞周期,未诱导HepG2与Jurkat共培养,凋亡1.4%,未见凋亡峰,加诱导的HepG2细胞株,Jurkat细胞的G1期变宽,G2期低平,G1峰前出现反映凋亡的亚G1峰,凋亡14.6%; 加抗FasL单克隆抗体的特异阻断试验,凋亡率降至5.1%(图2)。将表达FasL的HepG2的培养上清液加于Jurkat细胞,也可诱导后者凋亡,凋亡率36.8%;抗Fas单克隆抗体诱导的凋亡率为24.4%;而对照组仅见本底凋亡,凋亡率5.2%。
三、讨论
本文结果表明,不表达FasL的HepG2细胞与高表达Fas的细胞毒性T细胞Jurkat系可共生,经甲氨喋呤诱导的HepG2表达FasL后,则对Jurkat有明显杀伤作用,用抗FasL单抗则可阻断这一过程。说明FasL在肿瘤细胞杀伤淋巴细胞过程中具有细胞毒的作用。
, 百拇医药
淋巴细胞和肿瘤细胞都可以表达FasL,相互作用如何,取决于二者对Fas介导凋亡的敏感程度。许多肿瘤Fas表达蛋白丢失[2], 即使有残余Fas表达的癌细胞也逐渐耐受FasL介导的杀伤[3]。原因可能是有些原癌基因如bcl-2、bcr-abl活化可部分或完全阻断Fas凋亡的下游瀑布[4];同时激活T细胞对FasL介导的凋亡有一个易感窗口期。或可以解释为肿瘤细胞通过下调Fas及其它机制逐渐耐受淋巴细胞诱导的凋亡并构成表达FasL,对作为效应细胞的淋巴细胞进行免疫反击。
表达FasL的HepG2培养上清液也可诱导Jurkat系凋亡, 表明FasL是以旁分泌方式作用。可能游离FasL可使癌旁Fas阳性组织凋亡,为肿瘤侵袭和扩散提供空间。
眼前房和睾丸大量表达FasL并杀死浸润的淋巴细胞,创造了一个免疫特赦区[5]。FasL阳性的肿瘤组织则利用FasL表达杀伤淋巴细胞,完成免疫逃避、实现免疫反击。肿瘤组织中大量激活T细胞有利于肿瘤治疗,如何阻断肿瘤细胞对淋巴细胞的杀伤将成为研究热点。
, 百拇医药
参考文献
1 Zhu YF, Luo KX, Zhang LX, et al. In situ investigation of CD95L/CD95 expression in chronic hepatitis B virus infections. J Gastroenterol, Hepatol, 1997, 12:S121.
2 Leithauser F, Dhein J, Mechtersheimer G, et al. Constructive and induced expression of APO-1, a new member of the nerve growth factor/tumor necrosis factor receptor super family, in normal and neoplastic cells. J Exp Med, 1993, 177:195-200.
, 百拇医药
3 Shima YN, Nishimoto A, Ogata Y, et al. Myeloma cells express Fas antigen/APO-1(CD95) but only some are sensitive to anti-Fas antibody resulting in apoptosis. Blood, 1995, 85:757-764.
4 Jaattela MM, Benedict M, Tewari JA, et al. Bcl-x and Bcl-2 inhibit TNF and Fas-induced apoptosis and activation of phospholipase A2 in breast carcinoma cells. Oncogene, 1995, 10:2297-2305.
5 Griffith TS, Brunner T, Fletcher SM, et al. Fas ligand induced apoptosis as a mechanism of immune privilege. Science, 1995, 270:1189-1192.
(收稿:1997-12-16 修回:1998-05-19), 百拇医药
单位:510515 广州,第一军医大学南方医院
关键词:
中华医学杂志980807 我们应用免疫组织化学法发现肝癌病人的癌细胞Fas配体(FasL)阳性而汇管区浸润淋巴细胞发生凋亡[1]。为探讨FasL在肿瘤免疫逃避中的作用,用抗癌药诱导肝癌细胞株HepG2表达FasL,再与Jurkat T淋巴细胞共培养,经流式细胞仪研究其相互作用。
一、材料和方法
1.药物诱导实验:HepG2是1979年建立的人肝癌细胞株,Jurkat为人T淋巴细胞白血病细胞株,高水平表达Fas并对FasL及抗Fas单克隆抗体诱导的凋亡敏感(中国协和医科大学肿瘤研究所张叔人教授惠赠)。于HepG2细胞株中分别加入0、10、50、100、200 μg/ml甲氨喋呤12~24小时,收集细胞及上清液。常规提取RNA,用反义链引物合成cDNA。引物FasL-5′ATGTTTCAGCTCTTCCACCTACA GAAGGA3′和FasLR5′-TGACCAGAGAGAGCTCAGATACGTTGAC3′;β-actin-5′TGACGGGGTCACCCACACTGTGCCCATCT A3′和β-actinR-5′TCCTTAATGTCA CGCTCACGATTTC3′。常规PCR步骤。阳性对照用FasL质粒(广州中山医科大学徐安龙教授惠赠)。
, 百拇医药
2.共培养试验:HepG2细胞贴壁后加甲氨喋呤,孵育24小时去除上清液,PBS洗3次;5×106/ml Jurkat加入10 μmol/L 5-溴-2-脱氧尿嘧啶(BrdU)预孵育24小时作内源性DNA标志。每瓶(25 cm2)HepG2加入2.5×106 Jurkat,孵育24小时。HepG2加2 μg/ml抗FasL单克隆抗体,37 ℃温育1小时再加入Jurkat,作为FasL/Fas介导凋亡的特异阻断试验。HepG2处理后洗3次再孵育24小时,收集上清加入Jurkat培养48小时,观察上清中FasL活性。5×106/ml Jurkat细胞加入100 ng/ml抗Fas单克隆抗体(IgG3),为Fas介导凋亡的对照实验。
3.凋亡细胞的流式细胞仪检测:收集培养细胞,加入鼠抗BrdU抗体50 μl,再加抗鼠IgG异硫氰基荧光素(FITC)50 μl, 37℃各孵育30分钟;然后加含1%RNA 酶的Tris-HCl缓冲液500 μl孵育30分钟,加碘化丙啶(PI)进行DNA染色上机分析。
, 百拇医药
二、结果
1.药物诱导HepG2表达FasL:见图1。
注:M:DNA分子量标志;1.FasL质粒PCR产物;2.未诱导的HepG2细胞;3~6:HepG2细胞
图1 HepG2细胞经甲氨喋呤诱导表达
FasL mRNA
A.未诱导的HepG2与Jurkat共培养,图中A、B为本底凋亡值;B.诱导后的HepG2杀伤Jurkat细胞:出现凋亡的亚G1峰,HepG2被抗癌药杀伤,Jurkat被HepG2杀伤;C.HepG2经氨甲喋呤诱导,抗FasL阻断,再植入Jurkat, Jurkat仅有本底凋亡,HepG2凋亡率下降图2 氨甲喋呤诱导表达FasL的HepG2细胞对Jurkat细胞的杀伤
, 百拇医药
2.共培养及凋亡检测:以BrdU预标记Jurkat(标记率95%)、将2种细胞标记FITC,除去FITC低值的HepG2,分析FITC高值Jurkat的细胞周期,未诱导HepG2与Jurkat共培养,凋亡1.4%,未见凋亡峰,加诱导的HepG2细胞株,Jurkat细胞的G1期变宽,G2期低平,G1峰前出现反映凋亡的亚G1峰,凋亡14.6%; 加抗FasL单克隆抗体的特异阻断试验,凋亡率降至5.1%(图2)。将表达FasL的HepG2的培养上清液加于Jurkat细胞,也可诱导后者凋亡,凋亡率36.8%;抗Fas单克隆抗体诱导的凋亡率为24.4%;而对照组仅见本底凋亡,凋亡率5.2%。
三、讨论
本文结果表明,不表达FasL的HepG2细胞与高表达Fas的细胞毒性T细胞Jurkat系可共生,经甲氨喋呤诱导的HepG2表达FasL后,则对Jurkat有明显杀伤作用,用抗FasL单抗则可阻断这一过程。说明FasL在肿瘤细胞杀伤淋巴细胞过程中具有细胞毒的作用。
, 百拇医药
淋巴细胞和肿瘤细胞都可以表达FasL,相互作用如何,取决于二者对Fas介导凋亡的敏感程度。许多肿瘤Fas表达蛋白丢失[2], 即使有残余Fas表达的癌细胞也逐渐耐受FasL介导的杀伤[3]。原因可能是有些原癌基因如bcl-2、bcr-abl活化可部分或完全阻断Fas凋亡的下游瀑布[4];同时激活T细胞对FasL介导的凋亡有一个易感窗口期。或可以解释为肿瘤细胞通过下调Fas及其它机制逐渐耐受淋巴细胞诱导的凋亡并构成表达FasL,对作为效应细胞的淋巴细胞进行免疫反击。
表达FasL的HepG2培养上清液也可诱导Jurkat系凋亡, 表明FasL是以旁分泌方式作用。可能游离FasL可使癌旁Fas阳性组织凋亡,为肿瘤侵袭和扩散提供空间。
眼前房和睾丸大量表达FasL并杀死浸润的淋巴细胞,创造了一个免疫特赦区[5]。FasL阳性的肿瘤组织则利用FasL表达杀伤淋巴细胞,完成免疫逃避、实现免疫反击。肿瘤组织中大量激活T细胞有利于肿瘤治疗,如何阻断肿瘤细胞对淋巴细胞的杀伤将成为研究热点。
, 百拇医药
参考文献
1 Zhu YF, Luo KX, Zhang LX, et al. In situ investigation of CD95L/CD95 expression in chronic hepatitis B virus infections. J Gastroenterol, Hepatol, 1997, 12:S121.
2 Leithauser F, Dhein J, Mechtersheimer G, et al. Constructive and induced expression of APO-1, a new member of the nerve growth factor/tumor necrosis factor receptor super family, in normal and neoplastic cells. J Exp Med, 1993, 177:195-200.
, 百拇医药
3 Shima YN, Nishimoto A, Ogata Y, et al. Myeloma cells express Fas antigen/APO-1(CD95) but only some are sensitive to anti-Fas antibody resulting in apoptosis. Blood, 1995, 85:757-764.
4 Jaattela MM, Benedict M, Tewari JA, et al. Bcl-x and Bcl-2 inhibit TNF and Fas-induced apoptosis and activation of phospholipase A2 in breast carcinoma cells. Oncogene, 1995, 10:2297-2305.
5 Griffith TS, Brunner T, Fletcher SM, et al. Fas ligand induced apoptosis as a mechanism of immune privilege. Science, 1995, 270:1189-1192.
(收稿:1997-12-16 修回:1998-05-19), 百拇医药