fasЛљвђгыbcl-2ЗДвхRNAзЊЕМЮИАЉФЭвЉЯИАћЕФвЉУєЩЯЕїаЇгІ
зїепЃКаЄ Бљ ЪБгРШЋ едбрЧя гШ К Эѕзєгг СѕЯмСс вќ ЗМ ЧЧЬЋЖЋ ЗЎДњУї
ЕЅЮЛЃК
ЙиМќДЪЃКвЉЮяЩИбЁЪдбщЃЌПЙжзСіЃЛжзСіЯИАћЃЌХрбјЕФЃЛЮИжзСі/ВЁРэбЇЃЛдАЉЛљвђЕААзжЪРрЃЛЯИАћЕђЭі
Transduction of fas gene or bclTransduction of fas gene or bcl-2 antisense RNA sensitizes cultured drug resistant gastric cancer cells to chemotherapeutic drugs
XIAO Bing, SHI Yong-Quan, ZHAO Yan-Qiu, YOU Han, WANG Zuo-You, LIU Xian-Ling, YIN Fang, QIAO Tai-Dong and FAN Dai-Ming
, http://www.100md.com
Department of Gastroenterology, Xijing Hospital, Fourth Military Medical University, XiЃЇan 710032, Shaanxi Province, China
Subject headings drug screening assays, antitumor; tumor cells, cultured; stomach neoplasms/pathology; proto-oncogene proteins; apoptosis; Fas gene; bcl-2 gene
Abstract
AIM To compare the expression level of fas gene and bcl-2 gene in gastric cancer cells SGC7901 and in gastric cancer MDR cells SGC7901/ VCR, to transduce fas cDNA and bcl-2 antisense nucleic acid into SGC7901/ VCR cells respectively, and to observe the expression of two genes in transfectants and non-transfectants as well as their drug sensitivity.
, АйФДвНвЉ
METHODS The eukaryotic expression vector pBK-fas and pDOR-anti bcl-2 was constructed, and then transfected into SGC7901/ VCR cells by lipofectamine, respectively. The expression of mRNA and protein in SGC7901/ VCR cells and SGC7901 cells and transfectants was detected using Northern blot and Western blot and the drug sensitivity of transfectants for VCR, CDDP and 5-FU was analysed with MTT assay.
RESULTS After gene transfection, 80 for fas or 120 for antisense bcl-2 drug-resistant clones were selected from 2ЁС105 cells, the transfection efficiency was 0.04% and 0.06%, respectively. Two clones, SGC7901 fas/ VCR cells and SGC7901 anti-bcl-2/ VCR cells, were randomly selected for further incubation. Hybridization results showed that the expression level of fas mRNA and protein in SGC7901/ VCR cells was much lower, but the expression level of bcl-2 mRNA and protein was higher than that in SGC7901 cells, the expression level of fas mRNA and protein in SGC7901 fas/ VCR cells was higher, and the expression level of bcl-2 mRNA and protein was lower in SGC7901 anti bcl-2/ VCR cells than that in non-transfectants. MTT assay showed that transfectants were more sensitive to VCR, CDDP and 5-FU than non-transfectants.
, АйФДвНвЉ
CONCLUSION bcl-2 gene displayed high expression and fas gene displayed low expression in drug resistant gastric cancer cells. Expression of bcl-2 protein was effectively blocked in SGC7901 anti bcl-2/ VCR cells by gene transfection, in contrast, the expression of fas mRNA and protein in SGC7901 fas/ VCR cells increased. Fas gene and bcl-2 antisense nucleic acid transfection sensitized drug resistant gastric cancer cells to chemotherapeutic drugs.
жаЙњЭМЪщзЪСЯЗжРрКХ R 979.1
, АйФДвНвЉ
еЊ вЊ
ФПЕФ БШНЯfasЛљвђКЭbcl-2ЛљвђдкЮИАЉФЭвЉЯИАћгыЗЧФЭвЉЯИАћБэДяЕФВювьЃЌНЋfasЛљвђКЭbcl-2ЗДвхКЫЫсЕМШыЮИАЉФЭвЉЯИАћЃЌВЂЗжЮізЊЕМЧАКѓЕФЯИАћдкmRNAгыЕААзЕФБэДяЫЎЦНЃЌбаОПзЊЕМжъгыЗЧзЊЕМжъЖдЛЏСЦвЉЮяУєИаадЕФЧјБ№.
ЗНЗЈ ВЩгУЗжзгПЫТЁММЪѕНЋfas ЛљвђКЭЗДвхbcl-2ЦЌЖЮЗжБ№ВхШыецКЫБэДядиЬхpBK-CMVКЭpDOR-SV40ЕФЖрПЫТЁПЫТЁЮЛЕужЎМфЃЌвджЌжЪЬхНщЕМЗЈНЋСНИіжизщБэДядиЬхЗжБ№зЊШОЪмЬхЯИАћSGC7901/ VCRЃЌG418ЩИбЁПЫТЁЯИАћЃЌNorthern blot, Western blotМьВтФЭвЉЯИАћгыЗЧФЭвЉЯИАћвдМАзЊЕМЯИАћжаfasЛљвђКЭbcl-2ЛљвђmRNAМАЦфЕААзЕФБэДяЃЌMTTЗЈвЉУєЪЕбщМьВтзЊЕМЯИАћгыЗЧзЊЕМЯИАћЖдVCRЁЂЫГВЌЁЂ5-FU ЕФУєИаад.
НсЙћ ецКЫБэДядиЬхpBK-fas cDNAКЭpDOR-bcl-2 cDNAзЊЕМЮИАЉФЭвЉЯИАћКѓЃЌЗжБ№Дг2ЁС105ЯИАћжаЩИбЁГіДѓдМ80КЭ120ИіПЙадПЫТЁЃЌзЊЕМТЪИїЮЊ0.4ЁыКЭ0.6ЁыЃЌЫцЛњИїЬєбЁ2ИіПЫТЁМЬајЩИбЁгыРЉдіХрбјЃЌОљЛёЕУСЫЮШЖЈЕФПЙадЯИАћЃЌЮвУЧНЋДЫУќУћЮЊSGC7901 fas/ VCR cellКЭSGC7901 anti bcl-2/ VCR cell. дгНЛНсЙћБэУїЃЌЮИАЉФЭвЉЯИАћSGC7901/ VCRгыЗЧФЭвЉЯИАћЯрБШЃЌbcl-2ЛљвђЕФБэДяУїЯдНЯИпЃЌЖјfasЛљвђБэДяМЋЦфЮЂШѕ. зЊЕМжъSGC7901 fas/ VCR cellЕФfas mRNAМАЦфЕААзЫЎЦНЕФБэДяЯджјИпгкЗЧзЊЕМжъЃЌSGC7901 anti bcl-2/ VCR cellжаbcl-2ЕААзЕФБэДяУїЯдЕЭгкЗЧзЊЕМжъ. 2жъзЊЕМЯИАћЖдVCRЁЂЫГВЌЁЂ5-FUЕФУєИаадУїЯдИпгкЗЧзЊЕМжъ.
, http://www.100md.com
НсТл ЮИАЉФЭвЉЯИАћгыЗЧФЭвЉЯИАћЯрБШЃЌbcl-2ЛљвђДІгкИпБэДязДЬЌЃЌЖјfasЛљвђДІгкМЋЦфЕЭШѕЕФБэДязДЬЌ. ЭЈЙ§жЌжЪЬхНщЕМЕФЛљвђзЊШОЃЌгааЇЕизшЖЯСЫbcl-2ЕААздкSGC7901 anti bcl-2/ VCR cellЕФБэДяЃЌУїЯддіЧПСЫfas mRNAМАЦфЕААздкSGC7901 fas/ VCR cellЕФБэДя. bcl-2ЗДвхКЫЫсКЭfasЛљвђзЊЕМЮИАЉФЭвЉЯИАћКѓЃЌЖдЛЏСЦвЉЮяЕФУєИаадУїЯддіМг.
0 в§бд
ЛЏСЦЪЧжЮСЦжзСіЕФжївЊЗНЗЈжЎвЛЃЌЕЋФПЧАЫљбЁгУЕФЛЏСЦвЉЮяКЭгУвЉЗНАИЃЌФбвдДяЕНТњвтЕФСЦаЇЃЌЦфживЊЕФдвђжЎвЛОЭЪЧжзСіЯИАћЖдПЙАЉвЉЮяВњЩњСЫФЭвЉЯжЯѓ. вЊЯыЬсИпЛЏСЦаЇЙћЃЌОЭБиаыНтОіЖрвЉФЭвЉЕФЮЪЬт. НќФъбаОПБэУїЃЌжзСіЯИАћЕђЭіаХЯЂЕФвжжЦЁЂЯИАћЪйУќЕФбгГЄЪЧВњЩњЖрвЉФЭвЉЕФЛњжЦжЎвЛ[1]. вђДЫЃЌгаШЫЬсГіЭЈЙ§геЕМЛђДйНјЯИАћЕђЭіРДФцзЊжзСіЯИАћЕФФЭвЉЯжЯѓ. ЮвУЧНЋДйНјЕђЭіЕФfasЛљвђКЭbcl-2ЗДвхRNAЕМШыЮИАЉФЭвЉЯИАћSGC7901/ VCRЃЌВЂЙлВьСЫФПЕФЛљвђдкзЊЕМжъЕФБэДяМАЗжЮіСЫзЊЕМжъЖдЛЏСЦвЉЮяЕФУєИаадЃЌПЩЭћЮЊФцзЊЮИАЉЯИАћЖрвЉФЭвЉЯжЯѓЕФбаОПЕьЖЈЛљДЁ.
, http://www.100md.com
1 ВФСЯКЭЗНЗЈ
1.1 ВФСЯ ЮИАЉФЭвЉЯИАћSGC7901/ VCRМАJM109ОњжъЮЊБОЫљБЃДцЕФЯИАћгыОњжжЃЌФцзЊТМВЁЖОдиЬхpDOR-SV40КЭжЪСЃБэДядиЬхpBK-CMVгЩЕкЫФОќвНДѓбЇЛљДЁВПЩњЛЏНЬбаЪвДоДѓЯщВЉЪПЛндљЃЌpBluescript bcl-2 cDNAЮЊБОЫљБЃДцжЪСЃЃЌpBluescript fas cDNAЮЊШеБОItohНЬЪкЛндљ. EcoRЂё, BamHЂёЃЌSalЂёЃЌXhoЂёЃЌCIPЃЌ RNase AЃЌ T4 DNAСЌНгУИЃЌPMSFЃЌAprotininЃЌОлБћЯЉѕЃАЗЃЌABCЪдМСКаЃЌЬНеыБъМЧЪдМСКаМАвьСђЧшЫсывЕШЪдМСОљЙКгкЛЊУРЩњЮяЙЄГЬЙЋЫОКЭPromegaЙЋЫО. LipofectamineРДдДгкGibco BRL. MTTЃЌЕААзУИkЃЌDEPC,MOPS,G418ЃЌЬЅХЃбЊЧхМАЯѕЫсЯЫЮЌФЄЮЊSigmaВњЦЗ. [ІС-32P]dATPЙКгкББОЉбЧЛдЙЋЫОЃЌв§ЮяКЭЙбКЫмеЫсЬНеыКЯГЩгкЩЯКЃSangonЩњЮяжЦЦЗЙЋЫО. ЪѓПЙШЫbcl-2ПЙЬхКЭЭУПЙШЫfasПЙЬхОљЙКгкББОЉЖЋЗНЙЋЫО.
, АйФДвНвЉ
1.2 ЗНЗЈ
1.2.1 жизщдиЬхЕФЙЙНЈгыМјЖЈ ИљОнзЪСЯЃЌ1.9kbЕФbcl-2 cDNAдкpBluesecript KSЕФEcoRЂёЮЛЕужЎМфЃЌ2.5kbЕФfas cDNAдкpBluesecript KSжЪСЃЕФXhoЂёЮЛЕужЎМф. ИљОнжизщецКЫБэДядиЬхКЭЗДвхКЫЫсЙЙНЈЕФЗНЗЈЃЌдкУИЧаЕчгОжЄЪЕдЪМжЪСЃКЌгаЭъећЕФfas cDNAКЭbcl-2 cDNAЛљДЁЩЯЃЌРћгУдиЬхКЭЛљвђађСажаЕФЮЛЕуЃЌбЁдёEcoRЂёКЭBamHЂёЯћЛЏpBluesecript-bcl-2 cDNAЃЌSalЂёКЭEcoRЂёЯћЛЏpBluesecript-fas cDNAЃЌЖГШкЗЈЛиЪе0.622 kbЕФbcl-2 cDNAЦЌЖЮКЭ1.83 kbЕФfas cDNAЃЌдкT4 DNA ligaseзїгУЯТЃЌЗжБ№гыОЯргІЕФЯожЦадФкЧаУИЯћЛЏЕФpDORдиЬхМАpBK-CMVдиЬхDNAСЌНг(16Ёц,20 h)ЃЌзЊЛЏJM109ОњжъжЦБИЕФИаЪмЬЌЯИАћЃЌГЃЙцХрбј. ЫцЛњЬєбЁAmpПЙадОњТфМАаЁСПжЦБИжизщжЪСЃЃЌЭЈЙ§УИЧаМАЧэжЌЬЧФ§НКЕчгОЃЌМјЖЈЦфСЌНгаЇЙћгые§ШЗад. ИљОнfas cDNAађСаЗжЖЮЩшМЦ3Ждв§ЮяЃЌЗжБ№ЖдpBK-fas cDNAжизщзгНјааГЃЙцPCRРЉдіЃЌВЂНЋPCRВњЮяПЫТЁжСВтађдиЬхЃЌЫцЛњв§ЮяЗЈШЋздЖЏВтађЃЌЗжЮіжизщдиЬхжаФПЕФЛљвђЕФЖСПђ.
, http://www.100md.com
1.2.2 ЛљвђзЊЕМгыПЫТЁЩИбЁ АДееlipofectamineНщЕМЗЈЃЌШЁ0.5 g/ LДПЛЏЕФpDOR-bcl-2КЭpBK-fas cDNAИї1 ІЬLЃЌгУЮобЊЧхЕФRPMI1640ЯЁЪЭГЩ100 ІЬLЃЌВЂгыlipofectamine 5 ІЬLЛьКЯЃЌЪвЮТОВжУ10minЃЌзЊШОСљПзАхжаДІгкЖдЪ§ЩњГЄЦкЕФSGC7901/ VCRЯИАћ(2ЁС105 cells/Пз). 24 hКѓЃЌG418(500 mg/ L)ЩИбЁзЊШОЯИАћ. ЭЌЪБЃЌЩшСЂpDORКЭpBKПедиЬхзЊЕМЯИАћвдМАЗЧзЊЕМЯИАћзїЮЊЖдее. ПЫТЁЯИАћаЮГЩКѓЃЌж№ИіМЦЪ§ЃЌЫцЛњЬєбЁПЫТЁЃЌДѓСПРЉдіПЙадЯИАћ.
1.2.3 Southern blotКЭNorthern blot ЪеМЏЩњГЄзДЬЌСМКУЕФ1ЁС107ЯИАћЃЌАДЯИАћСбНтЗЈКЭвЛВНЗЈЗжБ№ЬсШЁЛљвђзщDNAКЭзмRNA. ЛљвђзщDNAОEcoRЂёЯћЛЏКѓЃЌЦеЭЈЧэжЌЬЧЕчгО. ЯИАћзмRNAОШШБфадКѓЃЌМзШЉБфадФ§НКЕчгО. ецПеГщЮќЗЈзЊвЦDNAКЭRNAжСЯѕЫсЯЫЮЌФЄЩЯ. вдЫцЛњв§ЮяЕФЗНЗЈжЦБИЭЌЮЛЫи[ІС-32P]dATPБъМЧЕФЬНеыВЂНјаадЄдгНЛЁЂдгНЛМАздЖЏЯдгА.
, http://www.100md.com
1.2.4 Western blot вдШ§ШЅЮлМССбНтЯИАћЕФЗНЗЈжЦБИЯИАћФЄЕААзЃЌШЁ1/ 2НјааSDS-PAGEЗжРыЕААзЬѕДјЃЌПМТэЫЙССРЖЯдЩЋМАЭбЩЋЃЌзаЯИЗжБцЬѕДјЕФВювь. СэвЛАыФЄЕААзМгбљЕчгОКѓЃЌ100 VЕчзЊвЦЙ§вЙЃЌSDSБфадNCФЄЃЌГЃЙцABCЗЈНсКЯПЙЬхгыЯдЩЋ.
1.2.5 вЉЮяУєИаадЪЕбщ НЋзЊЕМЯИАћКЭЗЧзЊЕМЯИАћАД103ЁЋ104 cellsНгжжгк96ПзАхЃЌдк200 ІЬLКЌ100 mL/ LаЁХЃбЊЧхЕФRPMI1640ХрбјвКжаЃЌЗжБ№МгШыЫГВЌЁЂ5-FUЃЌVCRвбжЊСйДВбЊНЌИпЗхХЈЖШЕФ0.1БЖ(1 ІЬg,7 ІЬg,0.1 ІЬg)ЁЂ1.0БЖ(10 ІЬg,70 ІЬg,1 ІЬg)МА10БЖ(100 ІЬg,700 ІЬg,10 ІЬg)ЕФМССПЃЌХрбј3 dЃЌМгШыMTTШмвК(5 g/ L) 20 ІЬLМЬајЗѕг§4 hЃЌЦњЩЯЧхЃЌМгШы150 ІЬL DMSOЃЌеёЕД10min вдШмНтНсОЇЃЌУИСЊУтвпМьВтвЧЩЯбЁдё590nmВЈГЄВтЖЈИїПзA(ODжЕ).
, АйФДвНвЉ 2 НсЙћ
2.1 ецКЫБэДядиЬхЕФЙЙНЈ дЪМжЪСЃЗжБ№ОEcoRЂёКЭXhoЂёУИЯћЛЏЃЌЗжРыГі1.9 kbЕФbcl-2 cDNAКЭ2.5 kbЕФfas cDNA(ЭМ1)ЃЌжизщдиЬхpDOR-bcl-2ОBamHЂёКЭEcoRЂёЫЋУИЧаЃЌЛёЕУ0.622 kbЕФЗДЯђbcl-2ЦЌЖЮКЭ6.5 kbЕФдиЬхЃЌPBK-fasбєаджизщзгОEcoRЂёгыSalЂёЯћЛЏЃЌПЩМћ1.8 kbЕФfas cDNAЦЌЖЮКЭ4.5 kbЕФдиЬхDNA. PBK-fasбєадОњТфЮЊФЃАхЕФPCRВњЮяЕчгОНсЙћШчЭМ2ЫљЪОЃЌ3Ждв§ЮяЗжБ№РЉдіГі500 bp,850 bp,700 bpЕФЦЌЖЮЃЌЦфДѓаЁИїгыв§ЮяжЎМфЕФађСаГЄЖШЯрЗћ. PCRВњЮяВтађНсЙћЃЌОЙ§аЃЖдгыЗжЮіЃЌгыЮФЯзСаГіЕФађСаЭъШЋвЛжТЃЌУЛгаШЮКЮДэЮѓЕФЖСПђ.
2.2 зЊЕМжъЕФНЈСЂ ЛљвђзЊЕМКѓЃЌ2ЁС105зЊШОЯИАћОG418ЩИбЁ4 wkЁЋ5 wkЃЌПедиЬхКЭБэДядиЬхЙВзЊШОЕФЯИАћХрбјПзФкОљж№НЅГіЯжПЙадПЫТЁЃЌОМЦЫуЗЂЯжЃЌЮШЖЈПЫТЁЪ§дк120зѓгвЃЌзЊЕМТЪдк0.5ЁывдЩЯ. ЖјЗЧзЊЕМжъвдЭЌМССПЕФG418ЩИбЁ2 wkЃЌЯИАћМДвбЭъШЋЫРЭі. ЫцЛњИїЬєбЁ2ИіЩњГЄзДЬЌСМКУЕФбєадПЫТЁЃЌМЬајвдПЙадЩИбЁЕФЭЌЪБЃЌНјааРЉдіХрбј40 dЁЋ50 dЃЌНсЙћИїЩИбЁГівЛИіЮШЖЈЕФПЙадЯИАћжъЃЌМДЗжБ№ЮЊSGC7901fas/ VCR cellsКЭSGC7901 anti bcl-2/ VCR cells.
, АйФДвНвЉ
2.3 ФПЕФЛљвђЕФБэДя Northern blot ЯдгАМћЭМ3КЭЭМ4. вдfas cDNAЕЅСДЬНеыЕФдгНЛНсЙћЃЌЮИАЉЯИАћШЁ3ЁС106 cellsЕФзмRNAЕчгОзЊФЄдгНЛЃЌГЪЯжЮЂШѕЕФЯдгАЃЌЖјЮИАЉФЭвЉвдЯрЭЌЯИАћЪ§ЕФдгНЛЯдгАИќМгЮЂШѕ. bcl-2 cDNAЕЅСДЬНеыЕФдгНЛЃЌдкЮИАЉЯИАћКЭЮИАЉФЭвЉЯИАћОљгаКмЧПЕФаХКХЃЌгШЦфЪЧФЭвЉЯИАћБэЯжИќМгЭЛГі. fasзЊЕМжъ1ЁС106 cellsЕФзмRNAзЊФЄдгНЛЃЌОпгаКмЧПЕФfasдгНЛаХКХЃЌЧвЯИАћЪ§СПдіМгЃЌаХКХдіЧП. ЗДвхbcl-2зЊЕМжъвдЗДвхЬНеыдгНЛЪБЃЌЦфдгНЛаХКХУїЯдЧПгкЗЧзЊЕМжъЃЌЖјвде§ЯђЬНеыдгНЛЪБЃЌдгНЛЯдгАЧПЖШТдЕЭгкЗЧзЊЕМжъ. SDS-PAGEЯдЩЋКЭWestern blotНсЙћМћЭМ5ЃЌfasзЊЕМжъдкMr 36 000ЁЋ40 000ДІБШЗЧзЊЕМжъУїЯдЖрГіЯжвЛЬѕЕчгОДјЃЌЦфДѓаЁгыfasЕААзЯрЖдЗжзгСПЯрЗћ. ЗЧзЊЕМжъдкMr 25 000ДІгавЛЬѕНЯЮЊХЈКёЕФЯдЩЋЃЌЦфДѓаЁгыbcl-2ЕААзЯрЖдЗжзгжЪСПвЛжТЃЌЖјЗДвхbcl-2зЊЕМжъЮДМћДЫЬѕДј. fasзЊЕМжъКЭЗЧзЊЕМжъЗжБ№гыfasПЙЬхКЭbcl-2ПЙЬхЕФНсКЯЃЌдкЯргІЕФЮЛжУвВГіЯжСЫгЁМЃзХЩЋ.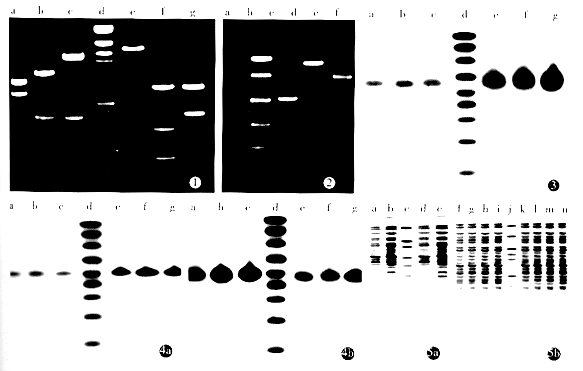
, АйФДвНвЉ
ЭМ1 жизщpBK-fasдиЬхКЭЗДвхКЫЫсpDOR-bcl-2ЕФУИЧаМјЖЈ
a.pBluescript fas/XhoЂё; b.pBluescript fas/EcoRЂё+SalЂё; c.pBK-fas/EcoRЂё+SalЂё; d.ІЫDNA marker/ HindЂѓ; e.pDOR bcl-2/ EcoRЂё+BamHЂё; f.pBluescript bcl-2/ EcoRЂё+BamHЂё; g.pBluescript bcl-2/ EcoRЂё.
ЭМ2 PBK-fas PCRВњЮяЕчгО
a.ИаЪмЬЌЯИАћЮЊФЃАхЕФPCR; b.ПедиЬхзЊЛЏОњжъЮЊФЃАхЕФPCR; c.PCR marker; d.pBK-fas PCRВњЮя1(500bp); e.pBK-fas PCR ВњЮя2(850bp); f.pBK-fas PCR ВњЮя3(700bp).
, http://www.100md.com
ЭМ3 Northern blot with fas cDNA probe
a.SGC7901/ VCR/ 3ЁС106 cells; b.ПедиЬхзЊЕМжъ/ 5ЁС106 cells; c.SGC7901/ 3ЁС106 cells; d.RNA marker; e.SGC7901 fas/ VCR/ 1ЁС106 cells; f.SGC7901 fas/ VCR/ 2ЁС106 cells; g.SGC7901 fas/ VCR/ 3ЁС106 cells.
ЭМ4 Northern blot with bcl-2 probe
A.bcl-2ЗДвхЬНеыЃЌ B.bcl-2е§вхЬНеы
a.SGC7901/ 3ЁС106 cells; b.ПедиЬхзЊЕМжъ/ 2ЁС106 cells; c.SGC7901/ 3ЁС106 cells; d.RNA marker; e.SGC7901 anti bcl-2/ VCR/ 2ЁС106 cells; f.SGC7901 anti bcl-2/ VCR/ 3ЁС106 cells; g.SGC7901 anti bcl-2/ VCR/ 4ЁС106 cells.
, АйФДвНвЉ
ЭМ5 SDS-PAGEКЭWestern blot
A.fasзЊЕМжъ B.bcl-2зЊЕМжъ
a.SGC7901/ VCR/ 3ЁС106 cells; b.SGC7901/ 3ЁС106 cells; c.ЕААзИпЗжзгСПБъзМВЮееЮя; d.SGC7901 fas/ VCR/ 1ЁС106 cells; e.SGC7901 fas/ VCR/ 2ЁС106 cells; f.SGC7901/ 1ЁС106 cells; g.SGC7901 / 2ЁС106 cells; h.SGC7901/ VCR/ 1ЁС106 cells; i.SGC7901/ VCR/ 2ЁС106 cells; j.ЕААзжаЗжзгСПБъзМВЮееЮя; k.SGC7901 anti bcl-2/ VCR/ 5ЁС105 cells; l.SGC7901 anti bcl-2/ VCR/ 1ЁС106 cells; m.SGC7901 anti bcl-2/ VCR/ 2ЁС106 cells; n.SGC7901 anti bcl-2/ VCR/ 3ЁС106 cells.
, АйФДвНвЉ
2.4 вЉЮяУєИаадЪЕбщНсЙћ ИљОнЯИАћДцЛюТЪ=(ЪЕбщзщA/ ЖдеезщA)ЁС100%ЃЌИїзщЯИАћЕФДцЛюТЪБШНЯШчБэ1ЫљЪОЃЌзЊЕМЯИАћОЫГВЌЁЂ5-FUзїгУЕФДцЛюТЪЃЌдк0.1БЖЁЂ1.0БЖМА10БЖЕФМССПОљУїЯдЕЭгкЗЧзЊЕМжъЃЌгШЦфЪЧfasЛљвђзЊЕМжъБэЯжИќМгУїЯд. зЊЕМЯИАћОVCRзїгУКѓЃЌгыЗЧзЊЕМжъЯрБШЃЌЫфШЛгавЛЖЈЕФНЕЕЭЃЌЕЋаЇЙћВЂВЛКмУїЯд.Бэ1 ЛЏСЦвЉЮязїгУгкзЊЕМжъгыЗЧзЊЕМжъЕФЯИАћДцЛюТЪ(%) (mg/ L)
Cells
n
CDDP
5-FU
VCR
5
50
, АйФДвНвЉ 500
35
350
3500
0.5
5
50
SGC7901 fas/ VCR
10
45.1a
40.5a
32.0
43.5a
, АйФДвНвЉ
38.6a
30.7a
55.0
46.7a
40.0a
SGC7901 anti bcl-2/ VCR
10
50.2a
45.1a
40.5
50.1a
, http://www.100md.com
44.6a
39.5a
60.3
55.1a
49.2a
SGC7901/ VCR
10
70.2
61.3
52.0
71.1
62.2
, http://www.100md.com
55.3
72.5
67.4
60.1
aP<0.05ЃЌ vs SGC7901/ VCR cells group.
3 ЬжТл
ЯИАћЕђЭіЪЧЩњЮяЬхвЛжжгыЯИАћгаЫПЗжСбЯрЗДЕФЗНЪНШЅЕїНкЯИАћШКЬхЯрЖдКтЖЈЕФживЊЗНЪН. НќФъШЯЮЊЃЌЯИАћЕђЭіЙ§ГЬЕФвжжЦЁЂЕђЭіЕїНкЛљвђЕФЮЩТвЃЌПЩЕМжТЯИАћЕђЭівжжЦЕААзЕФдіМгЛђЕђЭіДйНјвђзгЕФМѕЩйЃЌЪЧжзСіаЮГЩЕФСэвЛживЊЛњжЦ. ЭЈЙ§геЕМжзСіЯИАћЕФЕђЭіЃЌПЩвдДяЕНжЮСЦжзСіЕФФПЕФ. ФПЧАЪЙгУЕФаэЖрПЙАЉЛЏСЦвЉЮяжївЊЪЧЭЈЙ§вжжЦЛђЦЦЛЕЯИАћФкDNAКЯГЩвдМАЕїНкp53,c-myc,bcl-2,c-H-rasЕШЕђЭіЯрЙиЛљвђЃЌДЅЗЂЛђгеЕМЯИАћЕђЭіЃЌДгЖјвжжЦЛђЩБУ№жзСіЯИАћ[2-4]. ЕЋЪЧЃЌЛЏСЦвЉЮяВЛТлЭЈЙ§дѕбљЕФЛњжЦЦ№зїгУЃЌЖрвЉФЭвЉЕФЯжЯѓбЯжигАЯьСЫЖджзСіЕФжЮСЦаЇЙћ. НќФъРДЃЌЭЈЙ§ЩюШыбаОПЗЂЯжЃЌЕМжТжзСіЯИАћЗЂЩњЖрвЉФЭвЉЯжЯѓЕФЛњжЦЃЌГ§СЫPgpЃЌMRPЃЌLRPЃЌTopoЂђКЭGST/ GSHЕШФЭвЉЕААзКЭУИЕФИФБфжЎЭтЃЌЯИАћЕђЭіЕїПиЛњжЦЕФЮЩТввВИГгшжзСіЯИАћЖдЖржжНсЙЙЮоЙиЕФЛЏСЦвЉЮяВњЩњНЛВцПЙад[1ЃЌ5]. ФЭвЉЯИАћаоИДDNAЫ№ЩЫКЭЕжПЙЕђЭіЕФФмСІУїЯддіЧП[6]. ЯђЗЧФЭвЉЯИАћзЊЕМ bcl-xLЛљвђЃЌПЩгеЕМФЭвЉЯИАћБэаЭЕФГіЯжЃЌbcl-xLЛљвђЕФИпБэДягыPgpЕФИпБэДяГЪжБЯпЯрЙиЃЌВЂЖдаэЖрЛЏСЦвЉЮяВњЩњИпМССПЕФФЭЪмЃЌДгЖјШЯЮЊЕђЭіЯрЙиЛљвђЕФБэДязДЬЌОіЖЈСЫжзСіЯИАћЪЧЗёВњЩњФЭвЉЯжЯѓ[7]. ВжЪѓТбГВЯЫЮЌСіФЭвЉЯИАћLR73/ 20EгыЗЧФЭвЉЯИАћЯрБШЃЌдкИпБэДяMDRКЭPgpЕФЭЌЪБЃЌЖдЯИАћЕђЭігеЕММСcolchicineМЋВЛУєИаЃЌЕЋгУФЭвЉФцзЊМСverapamilДІРэФЭвЉЯИАћLR73/ 20EКѓЃЌУєИаадУїЯддіЧПЃЌДгЖјШЯЮЊФЭвЉЯИАћПЩозПЙЕђЭі[8]. ШщЯйАЉФЭвЉЯИАћMCF-7/ ADRИпБэДяbcl-2 mRNAМАЦфЕААзЃЌЭЈЙ§ЬЧМЂЖіЕФЗНЪНВЛФмгеЕМИУЯИАћЗЂЩњЕђЭі[9]. АзбЊВЁФЭвЉЯИАћHL-60/ ARЃЌHL-60/ VCRЃЌHL-60/ TAX1000гыЗЧФЭвЉЯИАћЯрБШЃЌдкИпБэДяPgpЃЌMRPЕФЭЌЪБЃЌвВИпБэДяbcl-2ЃЌbcl-xLЃЌВЂПЩвжжЦЛЏСЦвЉЮяpaclitaxelдкАћФкЕФОлМЏМАЦфгеЕМЕђЭіЕФзїгУ. ЗЧФЭвЉЯИАћзЊЕМbcl-2ЃЌbcl-xLЛљвђКѓВЛгАЯьЛЏСЦвЉЮяЕФАћФкОлМЏЃЌЕЋПЩвжжЦЯИАћЕђЭі[10]. fasПЙдКЭP53ЕААзЖМПЩвдДйНјЯИАћЕђЭіЃЌЕЋдкФЭвЉЯИАћжаЃЌfasКЭp53ЛљвђОљДІгкЕЭБэДязДЬЌ[1ЃЌ11]. ПЩМћЃЌbcl-2ЛљвђЕФИпБэДяМАfasЛљвђЕФЕЭБэДядкФЭвЉЯИАћозПЙЕђЭігыФЭЪмЛЏСЦвЉЮяЕФзїгУжаЃЌАчбнСЫживЊЕФНЧЩЋ. bcl-2ЛљвђЪЧЕђЭівжжЦЛљвђЃЌЫфШЛВЂВЛДйНјЯИАћЕФдіжГЃЌЕЋПЩУїЯдбгГЄЯИАћЕФДцЛюЪБМф. bcl-2ЭЈЙ§гАЯьАћФкЯИАћЦїЕФCa2+ЭЈЕРвдМАICEЕФБэДягыЪЭЗХЖјвжжЦЯИАћЕђЭі[12],ВЂгыBax,TNF,fasЕФНсКЯгыЕїНкгаЙи[13]. ЭЈЙ§ЗДвхКЫмеЫсММЪѕПижЦжзСіЯИАћФкbcl-2 mRNAЕФЗвыЃЌПЩгааЇвжжЦbcl-2ЛљвђЕФБэДяЃЌбгЛКжзСіЕФЩњГЄ[14].
, АйФДвНвЉ
fasЛљвђВњЮяЪЧвЛжжЕфаЭЕФЂёаЭФЄЕААзЃЌЪєTNFКЭNGFЪмЬхМвзхГЩдБ. ЯИАћБэУцfasЗжзггыЫќЕФХфЬхЗжзгfasLЛђПЙfasПЙЬхНсКЯЃЌПЩЯђЯИАћФкДЋЕнЫРЭіаХКХЃЌЪЙfasБэДяЯИАћдкЪ§аЁЪБФкЗЂЩњЫРЭі[15]. гыЯргІЕФе§ГЃзщжЏЯрБШЃЌЖёаджзСіЯИАћЯдЪОвьГЃЕЭЯТЕФfasБэДяЫЎЦНЃЌгШЦфЗЂЩњзЊвЦЕФжзСіМИКѕУЛгаБэДяЃЌЖјСМаджзСіЕФБэДяДѓЖрЪ§РрЫЦгкЦфРДдДЕФе§ГЃзщжЏ. гаШЫШЯЮЊЖёаджзСіЭЈЙ§ФГжжЛњжЦЃЌПЩФмвжжЦСЫfasЕФБэДяЛђНЋfasЖЊЪЇЃЌДгЖјЬгБмЛњЬхФкfasLЕФМрПи. дкФЭвЉЯИАћжаfasЛљвђЕФБэДяИќЮЊШБЗІ[1]. вђДЫЃЌЯђФЭвЉЯИАћФкзЊЕМПЩДйНјЯИАћЕђЭіЕФfasЛљвђЃЌЛђЭЈЙ§ЗДвхКЫЫсММЪѕзшЖЯФЭвЉЯИАћжаbcl-2ЛљвђЕФБэДяЃЌЮовЩгаРћгкдіЧПФЭвЉЯИАћЖдЛЏСЦвЉЮяЕФУєИаадЃЌДгЖјФцзЊФЭвЉЯжЯѓ. ЮвЙњбЇепГТаШКЕШШЫВЩгУЛљвђзЊШОММЪѕНЋbcl-2ЗДвхRNAЕМШыАзбЊВЁ(CEM)ЯИАћЃЌЙлВьСЫзувЖввпАЖдАаЯИАћЕФгАЯьЃЌНсЙћБэУїЃКЗДвхbcl-2ЛљвђЫВЪББэДяФмгааЇЕиНЕЕЭCEMФкдДадbcl-2ЕААзЫЎЦНЃЌВЂЪЙЦфЖдзувЖввпАЕФЯИАћЖОаЇгІИќЮЊУєИаЃЌЩБЩЫЯИАћАщгаДѓСПЯИАћЕђЭіаЁЬхгыЬнзДЭббѕКЫЬЧКЫЫсЦЌЖЮаЮГЩ.
, АйФДвНвЉ
ЮЊСЫУїШЗЮИАЉФЭвЉЯИАћЯЕSGC7901/ VCRжаfasгыbcl-2ЕААзЕФБэДяМАЬНЬжfasЛљвђКЭbcl-2ЗДвхКЫЫсзЊЕМSGC7901/ VCRЯИАћЕФжзСівжжЦгыФЭвЉФцзЊаЇгІЃЌЮвУЧВЩгУЗжзгПЫТЁММЪѕЃЌГЩЙІЕиЙЙНЈСЫецКЫБэДядиЬхpBK-fas cDNAгыpDOR-bcl-2 cDNA. ЭЈЙ§жЌжЪЬхНщЕМЕФзЊШОММЪѕЃЌНЋСНИіжизщБэДядиЬхЗжБ№зЊЕМФЭвЉЯИАћКѓЃЌЭЈЙ§ПЙадЩИбЁЃЌгааЇЕиНЈСЂСЫbcl-2зшЖЯЕФКЭfasЛљвђИпБэДяЕФЮИАЉФЭвЉЯИАћжъЃЌМДSGC7901 anti bcl-2/ VCR cellsКЭSGC7901 fas/ VCR cells. ЗжзгдгНЛНсЙћЯдЪОЃЌЮИАЉФЭвЉЯИАћSGC7901/ VCRгаМЋЦфЮЂСПЕФfas mRNAМАЦфЕААзБэДя. ЕЋbcl-2ГЪЯжИпБэДязДЬЌ. fasзЊЕМжъЕФfas mRNAМАЦфЕААзЕФБэДяУїЯдИпгкЗЧзЊЕМжъ. bcl-2зЊЕМжъжаЃЌbcl-2 mRNAЕФБэДяТдЮЂЪмЕНвжжЦЃЌЖјbcl-2ЕААзЫЎЦНЕФБэДяУїЯдЪмЕНзшЖЯ. вЉУєЪЕбщНсЙћБэУїЃЌfasЛљвђКЭЗДвхbcl-2зЊЕМЯИАћЖдЫГВЌЁЂ5-FUЕФУєИаадОљУїЯдИпгкЗЧзЊЕМжъЃЌЖдVCRЕФУєИаадвВгавЛЖЈЕФЬсИп. ДгЖјЫЕУїЃЌЯИАћЕђЭіЕФвжжЦдкжзСіЯИАћЖдЛЏСЦвЉЮяВњЩњФЭвЉЯжЯѓЕФЙ§ГЬжаОпгаживЊЕФзїгУЃЛгеЕМЯИАћЕђЭіЃЌдквЛЖЈГЬЖШЩЯПЩвдФцзЊетвЛФЭвЉЯжЯѓ.
, АйФДвНвЉ
ЕкЫФОќвНДѓбЇЮїОЉвНдКШЋОќЯћЛЏВЁбаОПЫљ ЩТЮїЪЁЮїАВЪа 710032
аЄБљЃЌФаЃЌ1962-09-11ЩњЃЌИЃНЈЪЁШ§УїЪаШЫЃЌККзх. 1996ФъЕквЛОќвНДѓбЇБЯвЕЃЌвНбЇВЉЪПЃЌЯћЛЏФкПЦжїжЮвНЪІЃЌбаОПЗНЯђЮЊжзСіЛљвђжЮСЦМАжзСіЯрЙиЛљвђЕФПЫТЁ.
ЭЈбЖзїеп ЗЎДњУїЃЌ710032ЃЌЕкЫФОќвНДѓбЇЮїОЉвНдКШЋОќЯћЛЏВЁбаОПЫљЃЌЩТЮїЪЁЮїАВЪаГЄРжЮїТЗ15КХ.
Correspondence to:FAN Dai-Ming, Department of Gastroenterology, Xijing Hospital, Fourth Military Medical University, XiЃЇan 710032, Shaanxi Province, China
Tel. +86ЁЄ29ЁЄ3232971-72863(O)
, АйФДвНвЉ
ЪеИхШеЦк 1998-06-06
4 ВЮПМЮФЯз
1 Friesen C, Fulda S, Debatin KM. Deficient activation of the CD95 (APO-1/ fas) system in drug-resistant cells. Leukemia, 1997;11(11):1833-1841
2 Arceci RJ. Tumor cell survival and resistance to therapy. Curr Opin Hematol, 1996;3(4):279-287
3 Ketley NJ, Allen PD, Kelsey SM, Newland AC. Modulation of idarubicin-induced apoptosis in human acute myeloid leukemia blasts by all-trans retinoic acid, 1,25(OH)2 vitamin D3, and granulocyte-macrophage colony-stimulating factor. Blood, 1997;90(11):4578-4587
, АйФДвНвЉ
4 Nooter K, Boersma AW, Oostrum RG, Burger H, Jochemsen AG, Stoter G. Constitutive expression of the c-H-ras oncogene inhibits doxorubicin-induced apoptosis and promotes cell survival in a rhabdomyosarcoma cell line. Br J Cancer, 1995;71(3):556-561
5 Van-Waardenburg RC, Meijer C, Burger H, Nooter K, De-Vries EG, Mulder NH et al. Effects of an inducible anti-sense c-myc gene transfer in a drug-resistant human small-cell-lung-carcinoma cell line. Int J Cancer, 1997;73(4):544-550
, АйФДвНвЉ
6 Colvin OM. Drug resistance in the treatment of sarcomas. Semin Oncol, 1997;24(5):580-591
7 Minn AJ, Rudin CM, Boise LH, Thompson CB. Expression of bcl-xL can confer a multidrug resistance phenotype. Blood, 1995;86(5):1903-1910
8 Robinson LJ, Roberts WK, Ling TT, Lamming D, Sternberg SS, Roepe PD. Human MDR 1 protein overexpression delays the apoptotic cascade in Chinese hamster ovary fibroblasts. Biochemistry, 1997;36(37):11169-11178
, http://www.100md.com
9 Lee YJ, Galoforo SS, Berns CM, Tong WP, Kim HR, Corry PM. Glucose deprivation-induced cytotoxicity in drug resistant human breast carcinoma MCF-7/ ADR cells: role of c-myc and bcl-2 in apoptotic cell death. J Cell Sci, 1997;110(Pt 5):681-686
10 Huang Y, Ibrado AM, Reed JC, Bullock G, Ray S, Tang C et al. Co-expression of several molecular mechanisms of multidrug resistance and their significance for paclitaxel cytotoxicity in human AML HL-60 cells. Leukemia, 1997;11(2):253-257
, http://www.100md.com
11 Reeve JG, xiong J, Morgan J, Bleehen NM. Expression of apoptosis-regulatory genes in lung tumour cell lines: relationship to p53 expression and relevance to acquired drug resistance. Br J Cancer, 1996;73(10):1193-1200
12 Shimizu S, Eguchi Y, Kamiikke W, Matsuda H, Tsujimoto Y. bcl-2 expression prevents activation of the ICE protease cascade. Oncogene, 1996;12(12):2251-2257
13 Allouche M, Bettaieb A, Vindis C, Rousse A, Grignon C, Laurent G. Influence of bcl-2 overexpression on the ceramide pathway in daunorubicin-induced apoptosis of leukemic cells. Oncogene, 1997;14(10):1837-1845
14 Webb A, Cunninngham D, Cotter F, Clarke PA, di-Stefano F, Ross P et al. bcl-2 antisense therapy in patients with non-Hodgkin lymphoma. Lancet, 1997;349(9059):1137-1141
15 Nagada S, Golstein P. The fas death factor. Science, 1995;267(5160):1449-1455, http://www.100md.com
ЕЅЮЛЃК
ЙиМќДЪЃКвЉЮяЩИбЁЪдбщЃЌПЙжзСіЃЛжзСіЯИАћЃЌХрбјЕФЃЛЮИжзСі/ВЁРэбЇЃЛдАЉЛљвђЕААзжЪРрЃЛЯИАћЕђЭі
Transduction of fas gene or bclTransduction of fas gene or bcl-2 antisense RNA sensitizes cultured drug resistant gastric cancer cells to chemotherapeutic drugs
XIAO Bing, SHI Yong-Quan, ZHAO Yan-Qiu, YOU Han, WANG Zuo-You, LIU Xian-Ling, YIN Fang, QIAO Tai-Dong and FAN Dai-Ming
, http://www.100md.com
Department of Gastroenterology, Xijing Hospital, Fourth Military Medical University, XiЃЇan 710032, Shaanxi Province, China
Subject headings drug screening assays, antitumor; tumor cells, cultured; stomach neoplasms/pathology; proto-oncogene proteins; apoptosis; Fas gene; bcl-2 gene
Abstract
AIM To compare the expression level of fas gene and bcl-2 gene in gastric cancer cells SGC7901 and in gastric cancer MDR cells SGC7901/ VCR, to transduce fas cDNA and bcl-2 antisense nucleic acid into SGC7901/ VCR cells respectively, and to observe the expression of two genes in transfectants and non-transfectants as well as their drug sensitivity.
, АйФДвНвЉ
METHODS The eukaryotic expression vector pBK-fas and pDOR-anti bcl-2 was constructed, and then transfected into SGC7901/ VCR cells by lipofectamine, respectively. The expression of mRNA and protein in SGC7901/ VCR cells and SGC7901 cells and transfectants was detected using Northern blot and Western blot and the drug sensitivity of transfectants for VCR, CDDP and 5-FU was analysed with MTT assay.
RESULTS After gene transfection, 80 for fas or 120 for antisense bcl-2 drug-resistant clones were selected from 2ЁС105 cells, the transfection efficiency was 0.04% and 0.06%, respectively. Two clones, SGC7901 fas/ VCR cells and SGC7901 anti-bcl-2/ VCR cells, were randomly selected for further incubation. Hybridization results showed that the expression level of fas mRNA and protein in SGC7901/ VCR cells was much lower, but the expression level of bcl-2 mRNA and protein was higher than that in SGC7901 cells, the expression level of fas mRNA and protein in SGC7901 fas/ VCR cells was higher, and the expression level of bcl-2 mRNA and protein was lower in SGC7901 anti bcl-2/ VCR cells than that in non-transfectants. MTT assay showed that transfectants were more sensitive to VCR, CDDP and 5-FU than non-transfectants.
, АйФДвНвЉ
CONCLUSION bcl-2 gene displayed high expression and fas gene displayed low expression in drug resistant gastric cancer cells. Expression of bcl-2 protein was effectively blocked in SGC7901 anti bcl-2/ VCR cells by gene transfection, in contrast, the expression of fas mRNA and protein in SGC7901 fas/ VCR cells increased. Fas gene and bcl-2 antisense nucleic acid transfection sensitized drug resistant gastric cancer cells to chemotherapeutic drugs.
жаЙњЭМЪщзЪСЯЗжРрКХ R 979.1
, АйФДвНвЉ
еЊ вЊ
ФПЕФ БШНЯfasЛљвђКЭbcl-2ЛљвђдкЮИАЉФЭвЉЯИАћгыЗЧФЭвЉЯИАћБэДяЕФВювьЃЌНЋfasЛљвђКЭbcl-2ЗДвхКЫЫсЕМШыЮИАЉФЭвЉЯИАћЃЌВЂЗжЮізЊЕМЧАКѓЕФЯИАћдкmRNAгыЕААзЕФБэДяЫЎЦНЃЌбаОПзЊЕМжъгыЗЧзЊЕМжъЖдЛЏСЦвЉЮяУєИаадЕФЧјБ№.
ЗНЗЈ ВЩгУЗжзгПЫТЁММЪѕНЋfas ЛљвђКЭЗДвхbcl-2ЦЌЖЮЗжБ№ВхШыецКЫБэДядиЬхpBK-CMVКЭpDOR-SV40ЕФЖрПЫТЁПЫТЁЮЛЕужЎМфЃЌвджЌжЪЬхНщЕМЗЈНЋСНИіжизщБэДядиЬхЗжБ№зЊШОЪмЬхЯИАћSGC7901/ VCRЃЌG418ЩИбЁПЫТЁЯИАћЃЌNorthern blot, Western blotМьВтФЭвЉЯИАћгыЗЧФЭвЉЯИАћвдМАзЊЕМЯИАћжаfasЛљвђКЭbcl-2ЛљвђmRNAМАЦфЕААзЕФБэДяЃЌMTTЗЈвЉУєЪЕбщМьВтзЊЕМЯИАћгыЗЧзЊЕМЯИАћЖдVCRЁЂЫГВЌЁЂ5-FU ЕФУєИаад.
НсЙћ ецКЫБэДядиЬхpBK-fas cDNAКЭpDOR-bcl-2 cDNAзЊЕМЮИАЉФЭвЉЯИАћКѓЃЌЗжБ№Дг2ЁС105ЯИАћжаЩИбЁГіДѓдМ80КЭ120ИіПЙадПЫТЁЃЌзЊЕМТЪИїЮЊ0.4ЁыКЭ0.6ЁыЃЌЫцЛњИїЬєбЁ2ИіПЫТЁМЬајЩИбЁгыРЉдіХрбјЃЌОљЛёЕУСЫЮШЖЈЕФПЙадЯИАћЃЌЮвУЧНЋДЫУќУћЮЊSGC7901 fas/ VCR cellКЭSGC7901 anti bcl-2/ VCR cell. дгНЛНсЙћБэУїЃЌЮИАЉФЭвЉЯИАћSGC7901/ VCRгыЗЧФЭвЉЯИАћЯрБШЃЌbcl-2ЛљвђЕФБэДяУїЯдНЯИпЃЌЖјfasЛљвђБэДяМЋЦфЮЂШѕ. зЊЕМжъSGC7901 fas/ VCR cellЕФfas mRNAМАЦфЕААзЫЎЦНЕФБэДяЯджјИпгкЗЧзЊЕМжъЃЌSGC7901 anti bcl-2/ VCR cellжаbcl-2ЕААзЕФБэДяУїЯдЕЭгкЗЧзЊЕМжъ. 2жъзЊЕМЯИАћЖдVCRЁЂЫГВЌЁЂ5-FUЕФУєИаадУїЯдИпгкЗЧзЊЕМжъ.
, http://www.100md.com
НсТл ЮИАЉФЭвЉЯИАћгыЗЧФЭвЉЯИАћЯрБШЃЌbcl-2ЛљвђДІгкИпБэДязДЬЌЃЌЖјfasЛљвђДІгкМЋЦфЕЭШѕЕФБэДязДЬЌ. ЭЈЙ§жЌжЪЬхНщЕМЕФЛљвђзЊШОЃЌгааЇЕизшЖЯСЫbcl-2ЕААздкSGC7901 anti bcl-2/ VCR cellЕФБэДяЃЌУїЯддіЧПСЫfas mRNAМАЦфЕААздкSGC7901 fas/ VCR cellЕФБэДя. bcl-2ЗДвхКЫЫсКЭfasЛљвђзЊЕМЮИАЉФЭвЉЯИАћКѓЃЌЖдЛЏСЦвЉЮяЕФУєИаадУїЯддіМг.
0 в§бд
ЛЏСЦЪЧжЮСЦжзСіЕФжївЊЗНЗЈжЎвЛЃЌЕЋФПЧАЫљбЁгУЕФЛЏСЦвЉЮяКЭгУвЉЗНАИЃЌФбвдДяЕНТњвтЕФСЦаЇЃЌЦфживЊЕФдвђжЎвЛОЭЪЧжзСіЯИАћЖдПЙАЉвЉЮяВњЩњСЫФЭвЉЯжЯѓ. вЊЯыЬсИпЛЏСЦаЇЙћЃЌОЭБиаыНтОіЖрвЉФЭвЉЕФЮЪЬт. НќФъбаОПБэУїЃЌжзСіЯИАћЕђЭіаХЯЂЕФвжжЦЁЂЯИАћЪйУќЕФбгГЄЪЧВњЩњЖрвЉФЭвЉЕФЛњжЦжЎвЛ[1]. вђДЫЃЌгаШЫЬсГіЭЈЙ§геЕМЛђДйНјЯИАћЕђЭіРДФцзЊжзСіЯИАћЕФФЭвЉЯжЯѓ. ЮвУЧНЋДйНјЕђЭіЕФfasЛљвђКЭbcl-2ЗДвхRNAЕМШыЮИАЉФЭвЉЯИАћSGC7901/ VCRЃЌВЂЙлВьСЫФПЕФЛљвђдкзЊЕМжъЕФБэДяМАЗжЮіСЫзЊЕМжъЖдЛЏСЦвЉЮяЕФУєИаадЃЌПЩЭћЮЊФцзЊЮИАЉЯИАћЖрвЉФЭвЉЯжЯѓЕФбаОПЕьЖЈЛљДЁ.
, http://www.100md.com
1 ВФСЯКЭЗНЗЈ
1.1 ВФСЯ ЮИАЉФЭвЉЯИАћSGC7901/ VCRМАJM109ОњжъЮЊБОЫљБЃДцЕФЯИАћгыОњжжЃЌФцзЊТМВЁЖОдиЬхpDOR-SV40КЭжЪСЃБэДядиЬхpBK-CMVгЩЕкЫФОќвНДѓбЇЛљДЁВПЩњЛЏНЬбаЪвДоДѓЯщВЉЪПЛндљЃЌpBluescript bcl-2 cDNAЮЊБОЫљБЃДцжЪСЃЃЌpBluescript fas cDNAЮЊШеБОItohНЬЪкЛндљ. EcoRЂё, BamHЂёЃЌSalЂёЃЌXhoЂёЃЌCIPЃЌ RNase AЃЌ T4 DNAСЌНгУИЃЌPMSFЃЌAprotininЃЌОлБћЯЉѕЃАЗЃЌABCЪдМСКаЃЌЬНеыБъМЧЪдМСКаМАвьСђЧшЫсывЕШЪдМСОљЙКгкЛЊУРЩњЮяЙЄГЬЙЋЫОКЭPromegaЙЋЫО. LipofectamineРДдДгкGibco BRL. MTTЃЌЕААзУИkЃЌDEPC,MOPS,G418ЃЌЬЅХЃбЊЧхМАЯѕЫсЯЫЮЌФЄЮЊSigmaВњЦЗ. [ІС-32P]dATPЙКгкББОЉбЧЛдЙЋЫОЃЌв§ЮяКЭЙбКЫмеЫсЬНеыКЯГЩгкЩЯКЃSangonЩњЮяжЦЦЗЙЋЫО. ЪѓПЙШЫbcl-2ПЙЬхКЭЭУПЙШЫfasПЙЬхОљЙКгкББОЉЖЋЗНЙЋЫО.
, АйФДвНвЉ
1.2 ЗНЗЈ
1.2.1 жизщдиЬхЕФЙЙНЈгыМјЖЈ ИљОнзЪСЯЃЌ1.9kbЕФbcl-2 cDNAдкpBluesecript KSЕФEcoRЂёЮЛЕужЎМфЃЌ2.5kbЕФfas cDNAдкpBluesecript KSжЪСЃЕФXhoЂёЮЛЕужЎМф. ИљОнжизщецКЫБэДядиЬхКЭЗДвхКЫЫсЙЙНЈЕФЗНЗЈЃЌдкУИЧаЕчгОжЄЪЕдЪМжЪСЃКЌгаЭъећЕФfas cDNAКЭbcl-2 cDNAЛљДЁЩЯЃЌРћгУдиЬхКЭЛљвђађСажаЕФЮЛЕуЃЌбЁдёEcoRЂёКЭBamHЂёЯћЛЏpBluesecript-bcl-2 cDNAЃЌSalЂёКЭEcoRЂёЯћЛЏpBluesecript-fas cDNAЃЌЖГШкЗЈЛиЪе0.622 kbЕФbcl-2 cDNAЦЌЖЮКЭ1.83 kbЕФfas cDNAЃЌдкT4 DNA ligaseзїгУЯТЃЌЗжБ№гыОЯргІЕФЯожЦадФкЧаУИЯћЛЏЕФpDORдиЬхМАpBK-CMVдиЬхDNAСЌНг(16Ёц,20 h)ЃЌзЊЛЏJM109ОњжъжЦБИЕФИаЪмЬЌЯИАћЃЌГЃЙцХрбј. ЫцЛњЬєбЁAmpПЙадОњТфМАаЁСПжЦБИжизщжЪСЃЃЌЭЈЙ§УИЧаМАЧэжЌЬЧФ§НКЕчгОЃЌМјЖЈЦфСЌНгаЇЙћгые§ШЗад. ИљОнfas cDNAађСаЗжЖЮЩшМЦ3Ждв§ЮяЃЌЗжБ№ЖдpBK-fas cDNAжизщзгНјааГЃЙцPCRРЉдіЃЌВЂНЋPCRВњЮяПЫТЁжСВтађдиЬхЃЌЫцЛњв§ЮяЗЈШЋздЖЏВтађЃЌЗжЮіжизщдиЬхжаФПЕФЛљвђЕФЖСПђ.
, http://www.100md.com
1.2.2 ЛљвђзЊЕМгыПЫТЁЩИбЁ АДееlipofectamineНщЕМЗЈЃЌШЁ0.5 g/ LДПЛЏЕФpDOR-bcl-2КЭpBK-fas cDNAИї1 ІЬLЃЌгУЮобЊЧхЕФRPMI1640ЯЁЪЭГЩ100 ІЬLЃЌВЂгыlipofectamine 5 ІЬLЛьКЯЃЌЪвЮТОВжУ10minЃЌзЊШОСљПзАхжаДІгкЖдЪ§ЩњГЄЦкЕФSGC7901/ VCRЯИАћ(2ЁС105 cells/Пз). 24 hКѓЃЌG418(500 mg/ L)ЩИбЁзЊШОЯИАћ. ЭЌЪБЃЌЩшСЂpDORКЭpBKПедиЬхзЊЕМЯИАћвдМАЗЧзЊЕМЯИАћзїЮЊЖдее. ПЫТЁЯИАћаЮГЩКѓЃЌж№ИіМЦЪ§ЃЌЫцЛњЬєбЁПЫТЁЃЌДѓСПРЉдіПЙадЯИАћ.
1.2.3 Southern blotКЭNorthern blot ЪеМЏЩњГЄзДЬЌСМКУЕФ1ЁС107ЯИАћЃЌАДЯИАћСбНтЗЈКЭвЛВНЗЈЗжБ№ЬсШЁЛљвђзщDNAКЭзмRNA. ЛљвђзщDNAОEcoRЂёЯћЛЏКѓЃЌЦеЭЈЧэжЌЬЧЕчгО. ЯИАћзмRNAОШШБфадКѓЃЌМзШЉБфадФ§НКЕчгО. ецПеГщЮќЗЈзЊвЦDNAКЭRNAжСЯѕЫсЯЫЮЌФЄЩЯ. вдЫцЛњв§ЮяЕФЗНЗЈжЦБИЭЌЮЛЫи[ІС-32P]dATPБъМЧЕФЬНеыВЂНјаадЄдгНЛЁЂдгНЛМАздЖЏЯдгА.
, http://www.100md.com
1.2.4 Western blot вдШ§ШЅЮлМССбНтЯИАћЕФЗНЗЈжЦБИЯИАћФЄЕААзЃЌШЁ1/ 2НјааSDS-PAGEЗжРыЕААзЬѕДјЃЌПМТэЫЙССРЖЯдЩЋМАЭбЩЋЃЌзаЯИЗжБцЬѕДјЕФВювь. СэвЛАыФЄЕААзМгбљЕчгОКѓЃЌ100 VЕчзЊвЦЙ§вЙЃЌSDSБфадNCФЄЃЌГЃЙцABCЗЈНсКЯПЙЬхгыЯдЩЋ.
1.2.5 вЉЮяУєИаадЪЕбщ НЋзЊЕМЯИАћКЭЗЧзЊЕМЯИАћАД103ЁЋ104 cellsНгжжгк96ПзАхЃЌдк200 ІЬLКЌ100 mL/ LаЁХЃбЊЧхЕФRPMI1640ХрбјвКжаЃЌЗжБ№МгШыЫГВЌЁЂ5-FUЃЌVCRвбжЊСйДВбЊНЌИпЗхХЈЖШЕФ0.1БЖ(1 ІЬg,7 ІЬg,0.1 ІЬg)ЁЂ1.0БЖ(10 ІЬg,70 ІЬg,1 ІЬg)МА10БЖ(100 ІЬg,700 ІЬg,10 ІЬg)ЕФМССПЃЌХрбј3 dЃЌМгШыMTTШмвК(5 g/ L) 20 ІЬLМЬајЗѕг§4 hЃЌЦњЩЯЧхЃЌМгШы150 ІЬL DMSOЃЌеёЕД10min вдШмНтНсОЇЃЌУИСЊУтвпМьВтвЧЩЯбЁдё590nmВЈГЄВтЖЈИїПзA(ODжЕ).
, АйФДвНвЉ 2 НсЙћ
2.1 ецКЫБэДядиЬхЕФЙЙНЈ дЪМжЪСЃЗжБ№ОEcoRЂёКЭXhoЂёУИЯћЛЏЃЌЗжРыГі1.9 kbЕФbcl-2 cDNAКЭ2.5 kbЕФfas cDNA(ЭМ1)ЃЌжизщдиЬхpDOR-bcl-2ОBamHЂёКЭEcoRЂёЫЋУИЧаЃЌЛёЕУ0.622 kbЕФЗДЯђbcl-2ЦЌЖЮКЭ6.5 kbЕФдиЬхЃЌPBK-fasбєаджизщзгОEcoRЂёгыSalЂёЯћЛЏЃЌПЩМћ1.8 kbЕФfas cDNAЦЌЖЮКЭ4.5 kbЕФдиЬхDNA. PBK-fasбєадОњТфЮЊФЃАхЕФPCRВњЮяЕчгОНсЙћШчЭМ2ЫљЪОЃЌ3Ждв§ЮяЗжБ№РЉдіГі500 bp,850 bp,700 bpЕФЦЌЖЮЃЌЦфДѓаЁИїгыв§ЮяжЎМфЕФађСаГЄЖШЯрЗћ. PCRВњЮяВтађНсЙћЃЌОЙ§аЃЖдгыЗжЮіЃЌгыЮФЯзСаГіЕФађСаЭъШЋвЛжТЃЌУЛгаШЮКЮДэЮѓЕФЖСПђ.
2.2 зЊЕМжъЕФНЈСЂ ЛљвђзЊЕМКѓЃЌ2ЁС105зЊШОЯИАћОG418ЩИбЁ4 wkЁЋ5 wkЃЌПедиЬхКЭБэДядиЬхЙВзЊШОЕФЯИАћХрбјПзФкОљж№НЅГіЯжПЙадПЫТЁЃЌОМЦЫуЗЂЯжЃЌЮШЖЈПЫТЁЪ§дк120зѓгвЃЌзЊЕМТЪдк0.5ЁывдЩЯ. ЖјЗЧзЊЕМжъвдЭЌМССПЕФG418ЩИбЁ2 wkЃЌЯИАћМДвбЭъШЋЫРЭі. ЫцЛњИїЬєбЁ2ИіЩњГЄзДЬЌСМКУЕФбєадПЫТЁЃЌМЬајвдПЙадЩИбЁЕФЭЌЪБЃЌНјааРЉдіХрбј40 dЁЋ50 dЃЌНсЙћИїЩИбЁГівЛИіЮШЖЈЕФПЙадЯИАћжъЃЌМДЗжБ№ЮЊSGC7901fas/ VCR cellsКЭSGC7901 anti bcl-2/ VCR cells.
, АйФДвНвЉ
2.3 ФПЕФЛљвђЕФБэДя Northern blot ЯдгАМћЭМ3КЭЭМ4. вдfas cDNAЕЅСДЬНеыЕФдгНЛНсЙћЃЌЮИАЉЯИАћШЁ3ЁС106 cellsЕФзмRNAЕчгОзЊФЄдгНЛЃЌГЪЯжЮЂШѕЕФЯдгАЃЌЖјЮИАЉФЭвЉвдЯрЭЌЯИАћЪ§ЕФдгНЛЯдгАИќМгЮЂШѕ. bcl-2 cDNAЕЅСДЬНеыЕФдгНЛЃЌдкЮИАЉЯИАћКЭЮИАЉФЭвЉЯИАћОљгаКмЧПЕФаХКХЃЌгШЦфЪЧФЭвЉЯИАћБэЯжИќМгЭЛГі. fasзЊЕМжъ1ЁС106 cellsЕФзмRNAзЊФЄдгНЛЃЌОпгаКмЧПЕФfasдгНЛаХКХЃЌЧвЯИАћЪ§СПдіМгЃЌаХКХдіЧП. ЗДвхbcl-2зЊЕМжъвдЗДвхЬНеыдгНЛЪБЃЌЦфдгНЛаХКХУїЯдЧПгкЗЧзЊЕМжъЃЌЖјвде§ЯђЬНеыдгНЛЪБЃЌдгНЛЯдгАЧПЖШТдЕЭгкЗЧзЊЕМжъ. SDS-PAGEЯдЩЋКЭWestern blotНсЙћМћЭМ5ЃЌfasзЊЕМжъдкMr 36 000ЁЋ40 000ДІБШЗЧзЊЕМжъУїЯдЖрГіЯжвЛЬѕЕчгОДјЃЌЦфДѓаЁгыfasЕААзЯрЖдЗжзгСПЯрЗћ. ЗЧзЊЕМжъдкMr 25 000ДІгавЛЬѕНЯЮЊХЈКёЕФЯдЩЋЃЌЦфДѓаЁгыbcl-2ЕААзЯрЖдЗжзгжЪСПвЛжТЃЌЖјЗДвхbcl-2зЊЕМжъЮДМћДЫЬѕДј. fasзЊЕМжъКЭЗЧзЊЕМжъЗжБ№гыfasПЙЬхКЭbcl-2ПЙЬхЕФНсКЯЃЌдкЯргІЕФЮЛжУвВГіЯжСЫгЁМЃзХЩЋ.
, АйФДвНвЉ
ЭМ1 жизщpBK-fasдиЬхКЭЗДвхКЫЫсpDOR-bcl-2ЕФУИЧаМјЖЈ
a.pBluescript fas/XhoЂё; b.pBluescript fas/EcoRЂё+SalЂё; c.pBK-fas/EcoRЂё+SalЂё; d.ІЫDNA marker/ HindЂѓ; e.pDOR bcl-2/ EcoRЂё+BamHЂё; f.pBluescript bcl-2/ EcoRЂё+BamHЂё; g.pBluescript bcl-2/ EcoRЂё.
ЭМ2 PBK-fas PCRВњЮяЕчгО
a.ИаЪмЬЌЯИАћЮЊФЃАхЕФPCR; b.ПедиЬхзЊЛЏОњжъЮЊФЃАхЕФPCR; c.PCR marker; d.pBK-fas PCRВњЮя1(500bp); e.pBK-fas PCR ВњЮя2(850bp); f.pBK-fas PCR ВњЮя3(700bp).
, http://www.100md.com
ЭМ3 Northern blot with fas cDNA probe
a.SGC7901/ VCR/ 3ЁС106 cells; b.ПедиЬхзЊЕМжъ/ 5ЁС106 cells; c.SGC7901/ 3ЁС106 cells; d.RNA marker; e.SGC7901 fas/ VCR/ 1ЁС106 cells; f.SGC7901 fas/ VCR/ 2ЁС106 cells; g.SGC7901 fas/ VCR/ 3ЁС106 cells.
ЭМ4 Northern blot with bcl-2 probe
A.bcl-2ЗДвхЬНеыЃЌ B.bcl-2е§вхЬНеы
a.SGC7901/ 3ЁС106 cells; b.ПедиЬхзЊЕМжъ/ 2ЁС106 cells; c.SGC7901/ 3ЁС106 cells; d.RNA marker; e.SGC7901 anti bcl-2/ VCR/ 2ЁС106 cells; f.SGC7901 anti bcl-2/ VCR/ 3ЁС106 cells; g.SGC7901 anti bcl-2/ VCR/ 4ЁС106 cells.
, АйФДвНвЉ
ЭМ5 SDS-PAGEКЭWestern blot
A.fasзЊЕМжъ B.bcl-2зЊЕМжъ
a.SGC7901/ VCR/ 3ЁС106 cells; b.SGC7901/ 3ЁС106 cells; c.ЕААзИпЗжзгСПБъзМВЮееЮя; d.SGC7901 fas/ VCR/ 1ЁС106 cells; e.SGC7901 fas/ VCR/ 2ЁС106 cells; f.SGC7901/ 1ЁС106 cells; g.SGC7901 / 2ЁС106 cells; h.SGC7901/ VCR/ 1ЁС106 cells; i.SGC7901/ VCR/ 2ЁС106 cells; j.ЕААзжаЗжзгСПБъзМВЮееЮя; k.SGC7901 anti bcl-2/ VCR/ 5ЁС105 cells; l.SGC7901 anti bcl-2/ VCR/ 1ЁС106 cells; m.SGC7901 anti bcl-2/ VCR/ 2ЁС106 cells; n.SGC7901 anti bcl-2/ VCR/ 3ЁС106 cells.
, АйФДвНвЉ
2.4 вЉЮяУєИаадЪЕбщНсЙћ ИљОнЯИАћДцЛюТЪ=(ЪЕбщзщA/ ЖдеезщA)ЁС100%ЃЌИїзщЯИАћЕФДцЛюТЪБШНЯШчБэ1ЫљЪОЃЌзЊЕМЯИАћОЫГВЌЁЂ5-FUзїгУЕФДцЛюТЪЃЌдк0.1БЖЁЂ1.0БЖМА10БЖЕФМССПОљУїЯдЕЭгкЗЧзЊЕМжъЃЌгШЦфЪЧfasЛљвђзЊЕМжъБэЯжИќМгУїЯд. зЊЕМЯИАћОVCRзїгУКѓЃЌгыЗЧзЊЕМжъЯрБШЃЌЫфШЛгавЛЖЈЕФНЕЕЭЃЌЕЋаЇЙћВЂВЛКмУїЯд.Бэ1 ЛЏСЦвЉЮязїгУгкзЊЕМжъгыЗЧзЊЕМжъЕФЯИАћДцЛюТЪ(%) (mg/ L)
Cells
n
CDDP
5-FU
VCR
5
50
, АйФДвНвЉ 500
35
350
3500
0.5
5
50
SGC7901 fas/ VCR
10
45.1a
40.5a
32.0
43.5a
, АйФДвНвЉ
38.6a
30.7a
55.0
46.7a
40.0a
SGC7901 anti bcl-2/ VCR
10
50.2a
45.1a
40.5
50.1a
, http://www.100md.com
44.6a
39.5a
60.3
55.1a
49.2a
SGC7901/ VCR
10
70.2
61.3
52.0
71.1
62.2
, http://www.100md.com
55.3
72.5
67.4
60.1
aP<0.05ЃЌ vs SGC7901/ VCR cells group.
3 ЬжТл
ЯИАћЕђЭіЪЧЩњЮяЬхвЛжжгыЯИАћгаЫПЗжСбЯрЗДЕФЗНЪНШЅЕїНкЯИАћШКЬхЯрЖдКтЖЈЕФживЊЗНЪН. НќФъШЯЮЊЃЌЯИАћЕђЭіЙ§ГЬЕФвжжЦЁЂЕђЭіЕїНкЛљвђЕФЮЩТвЃЌПЩЕМжТЯИАћЕђЭівжжЦЕААзЕФдіМгЛђЕђЭіДйНјвђзгЕФМѕЩйЃЌЪЧжзСіаЮГЩЕФСэвЛживЊЛњжЦ. ЭЈЙ§геЕМжзСіЯИАћЕФЕђЭіЃЌПЩвдДяЕНжЮСЦжзСіЕФФПЕФ. ФПЧАЪЙгУЕФаэЖрПЙАЉЛЏСЦвЉЮяжївЊЪЧЭЈЙ§вжжЦЛђЦЦЛЕЯИАћФкDNAКЯГЩвдМАЕїНкp53,c-myc,bcl-2,c-H-rasЕШЕђЭіЯрЙиЛљвђЃЌДЅЗЂЛђгеЕМЯИАћЕђЭіЃЌДгЖјвжжЦЛђЩБУ№жзСіЯИАћ[2-4]. ЕЋЪЧЃЌЛЏСЦвЉЮяВЛТлЭЈЙ§дѕбљЕФЛњжЦЦ№зїгУЃЌЖрвЉФЭвЉЕФЯжЯѓбЯжигАЯьСЫЖджзСіЕФжЮСЦаЇЙћ. НќФъРДЃЌЭЈЙ§ЩюШыбаОПЗЂЯжЃЌЕМжТжзСіЯИАћЗЂЩњЖрвЉФЭвЉЯжЯѓЕФЛњжЦЃЌГ§СЫPgpЃЌMRPЃЌLRPЃЌTopoЂђКЭGST/ GSHЕШФЭвЉЕААзКЭУИЕФИФБфжЎЭтЃЌЯИАћЕђЭіЕїПиЛњжЦЕФЮЩТввВИГгшжзСіЯИАћЖдЖржжНсЙЙЮоЙиЕФЛЏСЦвЉЮяВњЩњНЛВцПЙад[1ЃЌ5]. ФЭвЉЯИАћаоИДDNAЫ№ЩЫКЭЕжПЙЕђЭіЕФФмСІУїЯддіЧП[6]. ЯђЗЧФЭвЉЯИАћзЊЕМ bcl-xLЛљвђЃЌПЩгеЕМФЭвЉЯИАћБэаЭЕФГіЯжЃЌbcl-xLЛљвђЕФИпБэДягыPgpЕФИпБэДяГЪжБЯпЯрЙиЃЌВЂЖдаэЖрЛЏСЦвЉЮяВњЩњИпМССПЕФФЭЪмЃЌДгЖјШЯЮЊЕђЭіЯрЙиЛљвђЕФБэДязДЬЌОіЖЈСЫжзСіЯИАћЪЧЗёВњЩњФЭвЉЯжЯѓ[7]. ВжЪѓТбГВЯЫЮЌСіФЭвЉЯИАћLR73/ 20EгыЗЧФЭвЉЯИАћЯрБШЃЌдкИпБэДяMDRКЭPgpЕФЭЌЪБЃЌЖдЯИАћЕђЭігеЕММСcolchicineМЋВЛУєИаЃЌЕЋгУФЭвЉФцзЊМСverapamilДІРэФЭвЉЯИАћLR73/ 20EКѓЃЌУєИаадУїЯддіЧПЃЌДгЖјШЯЮЊФЭвЉЯИАћПЩозПЙЕђЭі[8]. ШщЯйАЉФЭвЉЯИАћMCF-7/ ADRИпБэДяbcl-2 mRNAМАЦфЕААзЃЌЭЈЙ§ЬЧМЂЖіЕФЗНЪНВЛФмгеЕМИУЯИАћЗЂЩњЕђЭі[9]. АзбЊВЁФЭвЉЯИАћHL-60/ ARЃЌHL-60/ VCRЃЌHL-60/ TAX1000гыЗЧФЭвЉЯИАћЯрБШЃЌдкИпБэДяPgpЃЌMRPЕФЭЌЪБЃЌвВИпБэДяbcl-2ЃЌbcl-xLЃЌВЂПЩвжжЦЛЏСЦвЉЮяpaclitaxelдкАћФкЕФОлМЏМАЦфгеЕМЕђЭіЕФзїгУ. ЗЧФЭвЉЯИАћзЊЕМbcl-2ЃЌbcl-xLЛљвђКѓВЛгАЯьЛЏСЦвЉЮяЕФАћФкОлМЏЃЌЕЋПЩвжжЦЯИАћЕђЭі[10]. fasПЙдКЭP53ЕААзЖМПЩвдДйНјЯИАћЕђЭіЃЌЕЋдкФЭвЉЯИАћжаЃЌfasКЭp53ЛљвђОљДІгкЕЭБэДязДЬЌ[1ЃЌ11]. ПЩМћЃЌbcl-2ЛљвђЕФИпБэДяМАfasЛљвђЕФЕЭБэДядкФЭвЉЯИАћозПЙЕђЭігыФЭЪмЛЏСЦвЉЮяЕФзїгУжаЃЌАчбнСЫживЊЕФНЧЩЋ. bcl-2ЛљвђЪЧЕђЭівжжЦЛљвђЃЌЫфШЛВЂВЛДйНјЯИАћЕФдіжГЃЌЕЋПЩУїЯдбгГЄЯИАћЕФДцЛюЪБМф. bcl-2ЭЈЙ§гАЯьАћФкЯИАћЦїЕФCa2+ЭЈЕРвдМАICEЕФБэДягыЪЭЗХЖјвжжЦЯИАћЕђЭі[12],ВЂгыBax,TNF,fasЕФНсКЯгыЕїНкгаЙи[13]. ЭЈЙ§ЗДвхКЫмеЫсММЪѕПижЦжзСіЯИАћФкbcl-2 mRNAЕФЗвыЃЌПЩгааЇвжжЦbcl-2ЛљвђЕФБэДяЃЌбгЛКжзСіЕФЩњГЄ[14].
, АйФДвНвЉ
fasЛљвђВњЮяЪЧвЛжжЕфаЭЕФЂёаЭФЄЕААзЃЌЪєTNFКЭNGFЪмЬхМвзхГЩдБ. ЯИАћБэУцfasЗжзггыЫќЕФХфЬхЗжзгfasLЛђПЙfasПЙЬхНсКЯЃЌПЩЯђЯИАћФкДЋЕнЫРЭіаХКХЃЌЪЙfasБэДяЯИАћдкЪ§аЁЪБФкЗЂЩњЫРЭі[15]. гыЯргІЕФе§ГЃзщжЏЯрБШЃЌЖёаджзСіЯИАћЯдЪОвьГЃЕЭЯТЕФfasБэДяЫЎЦНЃЌгШЦфЗЂЩњзЊвЦЕФжзСіМИКѕУЛгаБэДяЃЌЖјСМаджзСіЕФБэДяДѓЖрЪ§РрЫЦгкЦфРДдДЕФе§ГЃзщжЏ. гаШЫШЯЮЊЖёаджзСіЭЈЙ§ФГжжЛњжЦЃЌПЩФмвжжЦСЫfasЕФБэДяЛђНЋfasЖЊЪЇЃЌДгЖјЬгБмЛњЬхФкfasLЕФМрПи. дкФЭвЉЯИАћжаfasЛљвђЕФБэДяИќЮЊШБЗІ[1]. вђДЫЃЌЯђФЭвЉЯИАћФкзЊЕМПЩДйНјЯИАћЕђЭіЕФfasЛљвђЃЌЛђЭЈЙ§ЗДвхКЫЫсММЪѕзшЖЯФЭвЉЯИАћжаbcl-2ЛљвђЕФБэДяЃЌЮовЩгаРћгкдіЧПФЭвЉЯИАћЖдЛЏСЦвЉЮяЕФУєИаадЃЌДгЖјФцзЊФЭвЉЯжЯѓ. ЮвЙњбЇепГТаШКЕШШЫВЩгУЛљвђзЊШОММЪѕНЋbcl-2ЗДвхRNAЕМШыАзбЊВЁ(CEM)ЯИАћЃЌЙлВьСЫзувЖввпАЖдАаЯИАћЕФгАЯьЃЌНсЙћБэУїЃКЗДвхbcl-2ЛљвђЫВЪББэДяФмгааЇЕиНЕЕЭCEMФкдДадbcl-2ЕААзЫЎЦНЃЌВЂЪЙЦфЖдзувЖввпАЕФЯИАћЖОаЇгІИќЮЊУєИаЃЌЩБЩЫЯИАћАщгаДѓСПЯИАћЕђЭіаЁЬхгыЬнзДЭббѕКЫЬЧКЫЫсЦЌЖЮаЮГЩ.
, АйФДвНвЉ
ЮЊСЫУїШЗЮИАЉФЭвЉЯИАћЯЕSGC7901/ VCRжаfasгыbcl-2ЕААзЕФБэДяМАЬНЬжfasЛљвђКЭbcl-2ЗДвхКЫЫсзЊЕМSGC7901/ VCRЯИАћЕФжзСівжжЦгыФЭвЉФцзЊаЇгІЃЌЮвУЧВЩгУЗжзгПЫТЁММЪѕЃЌГЩЙІЕиЙЙНЈСЫецКЫБэДядиЬхpBK-fas cDNAгыpDOR-bcl-2 cDNA. ЭЈЙ§жЌжЪЬхНщЕМЕФзЊШОММЪѕЃЌНЋСНИіжизщБэДядиЬхЗжБ№зЊЕМФЭвЉЯИАћКѓЃЌЭЈЙ§ПЙадЩИбЁЃЌгааЇЕиНЈСЂСЫbcl-2зшЖЯЕФКЭfasЛљвђИпБэДяЕФЮИАЉФЭвЉЯИАћжъЃЌМДSGC7901 anti bcl-2/ VCR cellsКЭSGC7901 fas/ VCR cells. ЗжзгдгНЛНсЙћЯдЪОЃЌЮИАЉФЭвЉЯИАћSGC7901/ VCRгаМЋЦфЮЂСПЕФfas mRNAМАЦфЕААзБэДя. ЕЋbcl-2ГЪЯжИпБэДязДЬЌ. fasзЊЕМжъЕФfas mRNAМАЦфЕААзЕФБэДяУїЯдИпгкЗЧзЊЕМжъ. bcl-2зЊЕМжъжаЃЌbcl-2 mRNAЕФБэДяТдЮЂЪмЕНвжжЦЃЌЖјbcl-2ЕААзЫЎЦНЕФБэДяУїЯдЪмЕНзшЖЯ. вЉУєЪЕбщНсЙћБэУїЃЌfasЛљвђКЭЗДвхbcl-2зЊЕМЯИАћЖдЫГВЌЁЂ5-FUЕФУєИаадОљУїЯдИпгкЗЧзЊЕМжъЃЌЖдVCRЕФУєИаадвВгавЛЖЈЕФЬсИп. ДгЖјЫЕУїЃЌЯИАћЕђЭіЕФвжжЦдкжзСіЯИАћЖдЛЏСЦвЉЮяВњЩњФЭвЉЯжЯѓЕФЙ§ГЬжаОпгаживЊЕФзїгУЃЛгеЕМЯИАћЕђЭіЃЌдквЛЖЈГЬЖШЩЯПЩвдФцзЊетвЛФЭвЉЯжЯѓ.
, АйФДвНвЉ
ЕкЫФОќвНДѓбЇЮїОЉвНдКШЋОќЯћЛЏВЁбаОПЫљ ЩТЮїЪЁЮїАВЪа 710032
аЄБљЃЌФаЃЌ1962-09-11ЩњЃЌИЃНЈЪЁШ§УїЪаШЫЃЌККзх. 1996ФъЕквЛОќвНДѓбЇБЯвЕЃЌвНбЇВЉЪПЃЌЯћЛЏФкПЦжїжЮвНЪІЃЌбаОПЗНЯђЮЊжзСіЛљвђжЮСЦМАжзСіЯрЙиЛљвђЕФПЫТЁ.
ЭЈбЖзїеп ЗЎДњУїЃЌ710032ЃЌЕкЫФОќвНДѓбЇЮїОЉвНдКШЋОќЯћЛЏВЁбаОПЫљЃЌЩТЮїЪЁЮїАВЪаГЄРжЮїТЗ15КХ.
Correspondence to:FAN Dai-Ming, Department of Gastroenterology, Xijing Hospital, Fourth Military Medical University, XiЃЇan 710032, Shaanxi Province, China
Tel. +86ЁЄ29ЁЄ3232971-72863(O)
, АйФДвНвЉ
ЪеИхШеЦк 1998-06-06
4 ВЮПМЮФЯз
1 Friesen C, Fulda S, Debatin KM. Deficient activation of the CD95 (APO-1/ fas) system in drug-resistant cells. Leukemia, 1997;11(11):1833-1841
2 Arceci RJ. Tumor cell survival and resistance to therapy. Curr Opin Hematol, 1996;3(4):279-287
3 Ketley NJ, Allen PD, Kelsey SM, Newland AC. Modulation of idarubicin-induced apoptosis in human acute myeloid leukemia blasts by all-trans retinoic acid, 1,25(OH)2 vitamin D3, and granulocyte-macrophage colony-stimulating factor. Blood, 1997;90(11):4578-4587
, АйФДвНвЉ
4 Nooter K, Boersma AW, Oostrum RG, Burger H, Jochemsen AG, Stoter G. Constitutive expression of the c-H-ras oncogene inhibits doxorubicin-induced apoptosis and promotes cell survival in a rhabdomyosarcoma cell line. Br J Cancer, 1995;71(3):556-561
5 Van-Waardenburg RC, Meijer C, Burger H, Nooter K, De-Vries EG, Mulder NH et al. Effects of an inducible anti-sense c-myc gene transfer in a drug-resistant human small-cell-lung-carcinoma cell line. Int J Cancer, 1997;73(4):544-550
, АйФДвНвЉ
6 Colvin OM. Drug resistance in the treatment of sarcomas. Semin Oncol, 1997;24(5):580-591
7 Minn AJ, Rudin CM, Boise LH, Thompson CB. Expression of bcl-xL can confer a multidrug resistance phenotype. Blood, 1995;86(5):1903-1910
8 Robinson LJ, Roberts WK, Ling TT, Lamming D, Sternberg SS, Roepe PD. Human MDR 1 protein overexpression delays the apoptotic cascade in Chinese hamster ovary fibroblasts. Biochemistry, 1997;36(37):11169-11178
, http://www.100md.com
9 Lee YJ, Galoforo SS, Berns CM, Tong WP, Kim HR, Corry PM. Glucose deprivation-induced cytotoxicity in drug resistant human breast carcinoma MCF-7/ ADR cells: role of c-myc and bcl-2 in apoptotic cell death. J Cell Sci, 1997;110(Pt 5):681-686
10 Huang Y, Ibrado AM, Reed JC, Bullock G, Ray S, Tang C et al. Co-expression of several molecular mechanisms of multidrug resistance and their significance for paclitaxel cytotoxicity in human AML HL-60 cells. Leukemia, 1997;11(2):253-257
, http://www.100md.com
11 Reeve JG, xiong J, Morgan J, Bleehen NM. Expression of apoptosis-regulatory genes in lung tumour cell lines: relationship to p53 expression and relevance to acquired drug resistance. Br J Cancer, 1996;73(10):1193-1200
12 Shimizu S, Eguchi Y, Kamiikke W, Matsuda H, Tsujimoto Y. bcl-2 expression prevents activation of the ICE protease cascade. Oncogene, 1996;12(12):2251-2257
13 Allouche M, Bettaieb A, Vindis C, Rousse A, Grignon C, Laurent G. Influence of bcl-2 overexpression on the ceramide pathway in daunorubicin-induced apoptosis of leukemic cells. Oncogene, 1997;14(10):1837-1845
14 Webb A, Cunninngham D, Cotter F, Clarke PA, di-Stefano F, Ross P et al. bcl-2 antisense therapy in patients with non-Hodgkin lymphoma. Lancet, 1997;349(9059):1137-1141
15 Nagada S, Golstein P. The fas death factor. Science, 1995;267(5160):1449-1455, http://www.100md.com