不同胎龄新生儿免疫球蛋白重链可变区基因异型性
作者:肖昕 周晓光 熊爱华
单位:肖昕(510632 广州,暨南大学医学院附属第一医院儿科);熊爱华(510632 广州,暨南大学医学院附属第一医院药理教研室);周晓光(广东省妇幼保健院新生儿科)
关键词:
中华儿科杂志000519 免疫球蛋白重链可变区(immunoglobulin heavy chain variable region,IgH)是抗原特异性结合部位,在体液免疫中起关键作用[1,2]。编码人类IgH的胚系基因含有约200个可变区(VH)、20个多样化(D)和6个连接区(JH)基因片断[1]。IgH基因序列多样化是产生成千上万高特异性和高亲和力抗体的基础,它通过VH-D-JH连接的多样性和不精确性、N区(非基因编码的碱基序列)插入及VH基因片断上的体高突变等方式达到[1]。迄今为止,未见有人类不同胎龄新生儿IgH基因谱的系统分析和比较,故在本研究中,我们分析了极不成熟儿(25~30周)、不成熟儿(31~36周)和成熟儿(37~41周)IgH基因中的VH和JH基因片断的随机使用、体突变率、N区的插入和N区-D基因片段-N区(NDN)长度、开放性阅读框(open reading frame,ORF)等情况,以探讨新生儿成熟度对IgH基因多样性的影响。
, 百拇医药
对象和方法
已排除宫内细菌和病毒感染,胎龄分别为25~30周(10例)、31~36周(12例)和37~41周(11例)的新生儿,于出生时留取0.2 ml肝素抗凝脐血,用微量血DNA抽提试剂盒(QIAampTM,德国Qiagen公司)抽提DNA。应用PTC-100型DNA扩增仪(美国MJ research INS公司)对模板DNA进行2次扩增IgH基因序列(套式PCR,Nested PCR),扩增所用的第1套、第2套引物的碱基顺序分别为:5′CAGGTACAGCTGCA-GCAGTCAG 3′和5′TGAGGAGACGGTGACC 3′、5′TGGATCCG-CCAGGCTCTCNGG 3′和5′GTGACCAGGGTNCCTTGGCCCCAG 3′。 第1次扩增次数和条件为:在5个循环的变性94℃ 3 min、退火56℃ 3 min和延伸72℃ 2 min后,紧接着的是35个循环的变性94℃ 20 s、退火45℃ 30 s和延伸72℃ 2 min;第2次扩增次数和条件为:5个循环的变性94℃ 3 min、退火62℃ 3 min和延伸72℃ 2 min,然后是35个循环的变性94℃ 20 s、退火45℃ 30 s和延伸72℃ 1 min。用5%聚乙酰胺凝胶分离扩增物,第1次和第2次PCR产物均为多克隆性(在凝胶上呈云雾状条带形分布),其长度分别280~300和240~260 bp(图1)。应用LigATor plus Ligase试剂盒(德国R&D Systems 公司),将第2次扩增物与载体连结并转移到大肠杆菌内生长后,进行克隆筛选(每个新生儿挑选6个克隆)。根据Sanger的双脱氧末端终止法,使用ABI377型DNA序列自动分析仪(美国Perkin elmer公司)进行IgH基因序列测定。用电子计算机对所得到的序列与基因库中的IgH胚系基因序列及相互之间进行比较和分析,内容包括VH和JH基因片断的使用N区长度、体突变,以及开放性阅读框等。
, 百拇医药
统计学处理:采用统计软件SPSS for Windows 9.0处理,t检验、χ2检验或方差分析测试它们的差异显著性。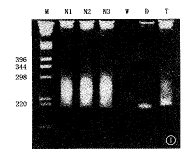
M:标志;N1:极不成熟儿;N2:不成熟儿;N3:成熟儿;W:水(阴性对照);D:Daudi DNA(阳性单克隆对照);T:扁桃体DNA(阳性多克隆对照)
图1 IgH基因第2次扩增产物电泳图(5%
聚乙酰胺凝胶,溴乙锭染色)
结果
极不成熟儿的60个克隆、不成熟儿的72个克隆和成熟儿的66个克隆被用于IgH基因序列分析,结果发现:
1.VH基因片段的使用率:极不成熟儿和不成熟儿优先使用VH1(40.0%和38.3%)和VH3(29.2%和33.3%);成熟儿主要使用VH3、VH5 和VH1(30.3%、21.2%和19.7%);随着胎龄增加,VH1和VH3的使用率逐渐下降(P<0.01),其余VH的使用率有所增加(P<0.05)。
, http://www.100md.com
2.JH基因片段使用率:在极不成熟儿和不成熟儿,优先使用JH3(46.7%和40.3%)和JH6(35.0%和41.7%),JH4的使用率(8.3%和6.9%)和JH5的使用率(均为8.3%)较低,JH2偶被使用(2.7%),JH1未被使用;在成熟儿,JH3的使用率相对较高(31.8%),JH6、JH5和JH4的使用率相似(22.7%、21.2%和21.2%),JH2的使用率仍不高(3.0%),JH1仍未被使用;随着胎龄增加,JH3和JH6的使用率下降(P<0.01),JH4和JH5的使用率上升(P<0.01),JH2的使用率变化不大(P>0.05)。
, http://www.100md.com
3.D基因片段的使用、N区的插入及NDN长度:在极不成熟儿、不成熟儿和成熟儿,均优先使用D2基因片段,使用率分别为45.0%、45.8%和48.6%,三者差异无显著性(P>0.05)。在IgH基因序列的VH-D-JH连接点处,极不成熟儿无N区插入,不成熟儿9.7%的序列有N区(1~5个碱基)插入,成熟儿21.2%的序列有数量不等的N区(1~9个碱基)插入;NDN长度分别为(13.5±7.2)、(16.8±6.5)和(22.3±7.2) bp,差异有非常显著性(P<0.01)。
4.H基因片段上的体突变:在极不成熟儿,13.3%的 IgH基因序列存在点突变(仅在1个位点);在不成熟儿,16.7%序列存在点突变(9个为1个位点的点突变,3个为2个位点的点突变);在成熟儿,30.3%序列存在点突变(12个为1个位点的点突变,8个为2个位点或以上的点突变)。上述的点突变分别属于A→G、C→G、T→C、G→T或C→T的置换突变。
, 百拇医药
5.ORF:在极不成熟儿、不成熟儿和成熟儿,分别65.0%、68.1%和92.4%的IgH基因序列有ORF,成熟儿与极不成熟儿或不成熟儿差异有非常显著性(P<0.01),而不成熟儿与极不成熟儿之间,差异无显著性(P>0.05)。尽管如此,ORF仍是随新生儿胎龄的增加而增加(P<0.05)。
讨论
动物实验表明,针对特异性抗原起反应的免疫系统的发育是一个受控的渐进过程,对此过程具有重要影响的事件就是IgH胚系基因的重排。它是在B淋巴细胞发育和成熟过程中,IgH胚系基因实现VH-D-JH连接后形成。IgH基因重排多样化使得抗体谱多样性增加[1,2]。本研究发现,胎龄25周以上的人类早产儿(包括极不成熟儿和不成熟儿)即有重排的IgH基因存在,加上他们能产生抗体的事实[3],说明人类新生儿的早期生长发育阶段,使VH-D-JH实现连接的机制已处于活化状态。这种处于活化状态的重排IgH基因是完整、具有功能的基因,能够转录mRNA并翻译免疫球蛋白(抗体)[1]。至于为何早产儿产生的抗体不但量少而且亲和力低,也可以用本研究的结果来解释:由于在极不成熟儿和不成熟儿的重排IgH基因中,VH、JH基因片断的随机使用率和内在性体突变率低、NDN长度短及ORF少等原因,基因多样性极其有限,故其编码的抗体谱多样性和亲和力较低。
, http://www.100md.com
动物实验结果提示:胎鼠和新生鼠的IgH重排基因的VH-D-JH连接点处的N区缺乏或虽有插入但极短;胎猪IgH基因重排中,某些VH基因片断被偏倚使用[4,5]。Cuisinier等[6]发现胎儿优先使用接近D基因片段的VH基因片段如VH5和VH6,Stollar[7]则发现成人VH、D和JH基因片段的使用是随机的,有较长的N区和高体突变存在。本研究首次发现,不同胎龄组新生儿IgH基因存在异型性:在早产儿,偏倚使用VH1和VH3,而在成熟儿(足月儿),则主要使用VH3 、VH5和 VH1。胎龄越小,NDN长度越短、体突变率越低,以及开放性阅读框越少。至于JH基因片段使用,在所有的新生儿,JH3的使用率均最高,JH2偶被使用,JH1未被使用;随着胎龄增加,JH3和JH6的使用率下降,JH4和JH5的使用率上升。在体突变的克隆中,大多数为置换突变A→G、C→G、T→C、G→T或C→T。综合Cuisinier、Choi和本研究的发现,可以推测:新生儿成熟度对IgH基因的重排产生明显影响,即随着新生儿的不断成熟,VH、D和JH基因片断的随机使用率,内在性体突变率、N区的长度,以及开放性阅读框等也增加,进而IgH基因多样性逐渐增加,表明人类新生儿和(或)胎儿体液免疫系统的发育也是一个渐进的发展过程。
, http://www.100md.com
基金项目:国家自然科学基金资助项目(39800175);广东省自然科学基金资助项目(980713)
参考文献
1,Tonegawa S. Somatic generation of antibody diversity. Nature, 1983, 302 :575-581.
2,Bhat NM, Bieber MM, Teng NN. Heavy chain variable gene usage by human B-1 lymphocytes and polyreactive autoantibodies. Hum Antibo, 1997, 8:146-150.
3,Ballow M, Cates KL, Rowe JC, et al. Development of the immune system in very low birth weight (less than 1 500g) premature infants: concentrations of plasma immunoglobulins and patterns of infections. Pediatr Res, 1986, 20:889-904.
, http://www.100md.com
4,Feeney AJ. Lack of N regions in fetal and neonatal mouse Ig V-D-J junctional sequences. J Exp Med, 1990, 172:1377-1390.
5,Sun J, Hayward C, Shinde R, et al. Antibody repertoire development in fetal and neonatal piglets. I. Four VH genes account for 80 percent of VH usage during 84 days of fetal life. J Immunol, 1998, 161:5070-5078.
6,Cuisinier AM, Guigou V, Boubli L, et al. Preferential expression of VH5 and VH6 immunoglobulin genes in early human B-cell ontogeny. Scand J Immunol, 1989, 30:493-497.
7,Stollar BD. The expressed heavy chain V gene repertorire of circulating B cells in normal adults. Ann N Y Acad Sci, 1995, 764:265-274.
(收稿日期:1999-07-08), 百拇医药
单位:肖昕(510632 广州,暨南大学医学院附属第一医院儿科);熊爱华(510632 广州,暨南大学医学院附属第一医院药理教研室);周晓光(广东省妇幼保健院新生儿科)
关键词:
中华儿科杂志000519 免疫球蛋白重链可变区(immunoglobulin heavy chain variable region,IgH)是抗原特异性结合部位,在体液免疫中起关键作用[1,2]。编码人类IgH的胚系基因含有约200个可变区(VH)、20个多样化(D)和6个连接区(JH)基因片断[1]。IgH基因序列多样化是产生成千上万高特异性和高亲和力抗体的基础,它通过VH-D-JH连接的多样性和不精确性、N区(非基因编码的碱基序列)插入及VH基因片断上的体高突变等方式达到[1]。迄今为止,未见有人类不同胎龄新生儿IgH基因谱的系统分析和比较,故在本研究中,我们分析了极不成熟儿(25~30周)、不成熟儿(31~36周)和成熟儿(37~41周)IgH基因中的VH和JH基因片断的随机使用、体突变率、N区的插入和N区-D基因片段-N区(NDN)长度、开放性阅读框(open reading frame,ORF)等情况,以探讨新生儿成熟度对IgH基因多样性的影响。
, 百拇医药
对象和方法
已排除宫内细菌和病毒感染,胎龄分别为25~30周(10例)、31~36周(12例)和37~41周(11例)的新生儿,于出生时留取0.2 ml肝素抗凝脐血,用微量血DNA抽提试剂盒(QIAampTM,德国Qiagen公司)抽提DNA。应用PTC-100型DNA扩增仪(美国MJ research INS公司)对模板DNA进行2次扩增IgH基因序列(套式PCR,Nested PCR),扩增所用的第1套、第2套引物的碱基顺序分别为:5′CAGGTACAGCTGCA-GCAGTCAG 3′和5′TGAGGAGACGGTGACC 3′、5′TGGATCCG-CCAGGCTCTCNGG 3′和5′GTGACCAGGGTNCCTTGGCCCCAG 3′。 第1次扩增次数和条件为:在5个循环的变性94℃ 3 min、退火56℃ 3 min和延伸72℃ 2 min后,紧接着的是35个循环的变性94℃ 20 s、退火45℃ 30 s和延伸72℃ 2 min;第2次扩增次数和条件为:5个循环的变性94℃ 3 min、退火62℃ 3 min和延伸72℃ 2 min,然后是35个循环的变性94℃ 20 s、退火45℃ 30 s和延伸72℃ 1 min。用5%聚乙酰胺凝胶分离扩增物,第1次和第2次PCR产物均为多克隆性(在凝胶上呈云雾状条带形分布),其长度分别280~300和240~260 bp(图1)。应用LigATor plus Ligase试剂盒(德国R&D Systems 公司),将第2次扩增物与载体连结并转移到大肠杆菌内生长后,进行克隆筛选(每个新生儿挑选6个克隆)。根据Sanger的双脱氧末端终止法,使用ABI377型DNA序列自动分析仪(美国Perkin elmer公司)进行IgH基因序列测定。用电子计算机对所得到的序列与基因库中的IgH胚系基因序列及相互之间进行比较和分析,内容包括VH和JH基因片断的使用N区长度、体突变,以及开放性阅读框等。
, 百拇医药
统计学处理:采用统计软件SPSS for Windows 9.0处理,t检验、χ2检验或方差分析测试它们的差异显著性。
M:标志;N1:极不成熟儿;N2:不成熟儿;N3:成熟儿;W:水(阴性对照);D:Daudi DNA(阳性单克隆对照);T:扁桃体DNA(阳性多克隆对照)
图1 IgH基因第2次扩增产物电泳图(5%
聚乙酰胺凝胶,溴乙锭染色)
结果
极不成熟儿的60个克隆、不成熟儿的72个克隆和成熟儿的66个克隆被用于IgH基因序列分析,结果发现:
1.VH基因片段的使用率:极不成熟儿和不成熟儿优先使用VH1(40.0%和38.3%)和VH3(29.2%和33.3%);成熟儿主要使用VH3、VH5 和VH1(30.3%、21.2%和19.7%);随着胎龄增加,VH1和VH3的使用率逐渐下降(P<0.01),其余VH的使用率有所增加(P<0.05)。
, http://www.100md.com
2.JH基因片段使用率:在极不成熟儿和不成熟儿,优先使用JH3(46.7%和40.3%)和JH6(35.0%和41.7%),JH4的使用率(8.3%和6.9%)和JH5的使用率(均为8.3%)较低,JH2偶被使用(2.7%),JH1未被使用;在成熟儿,JH3的使用率相对较高(31.8%),JH6、JH5和JH4的使用率相似(22.7%、21.2%和21.2%),JH2的使用率仍不高(3.0%),JH1仍未被使用;随着胎龄增加,JH3和JH6的使用率下降(P<0.01),JH4和JH5的使用率上升(P<0.01),JH2的使用率变化不大(P>0.05)。
, http://www.100md.com
3.D基因片段的使用、N区的插入及NDN长度:在极不成熟儿、不成熟儿和成熟儿,均优先使用D2基因片段,使用率分别为45.0%、45.8%和48.6%,三者差异无显著性(P>0.05)。在IgH基因序列的VH-D-JH连接点处,极不成熟儿无N区插入,不成熟儿9.7%的序列有N区(1~5个碱基)插入,成熟儿21.2%的序列有数量不等的N区(1~9个碱基)插入;NDN长度分别为(13.5±7.2)、(16.8±6.5)和(22.3±7.2) bp,差异有非常显著性(P<0.01)。
4.H基因片段上的体突变:在极不成熟儿,13.3%的 IgH基因序列存在点突变(仅在1个位点);在不成熟儿,16.7%序列存在点突变(9个为1个位点的点突变,3个为2个位点的点突变);在成熟儿,30.3%序列存在点突变(12个为1个位点的点突变,8个为2个位点或以上的点突变)。上述的点突变分别属于A→G、C→G、T→C、G→T或C→T的置换突变。
, 百拇医药
5.ORF:在极不成熟儿、不成熟儿和成熟儿,分别65.0%、68.1%和92.4%的IgH基因序列有ORF,成熟儿与极不成熟儿或不成熟儿差异有非常显著性(P<0.01),而不成熟儿与极不成熟儿之间,差异无显著性(P>0.05)。尽管如此,ORF仍是随新生儿胎龄的增加而增加(P<0.05)。
讨论
动物实验表明,针对特异性抗原起反应的免疫系统的发育是一个受控的渐进过程,对此过程具有重要影响的事件就是IgH胚系基因的重排。它是在B淋巴细胞发育和成熟过程中,IgH胚系基因实现VH-D-JH连接后形成。IgH基因重排多样化使得抗体谱多样性增加[1,2]。本研究发现,胎龄25周以上的人类早产儿(包括极不成熟儿和不成熟儿)即有重排的IgH基因存在,加上他们能产生抗体的事实[3],说明人类新生儿的早期生长发育阶段,使VH-D-JH实现连接的机制已处于活化状态。这种处于活化状态的重排IgH基因是完整、具有功能的基因,能够转录mRNA并翻译免疫球蛋白(抗体)[1]。至于为何早产儿产生的抗体不但量少而且亲和力低,也可以用本研究的结果来解释:由于在极不成熟儿和不成熟儿的重排IgH基因中,VH、JH基因片断的随机使用率和内在性体突变率低、NDN长度短及ORF少等原因,基因多样性极其有限,故其编码的抗体谱多样性和亲和力较低。
, http://www.100md.com
动物实验结果提示:胎鼠和新生鼠的IgH重排基因的VH-D-JH连接点处的N区缺乏或虽有插入但极短;胎猪IgH基因重排中,某些VH基因片断被偏倚使用[4,5]。Cuisinier等[6]发现胎儿优先使用接近D基因片段的VH基因片段如VH5和VH6,Stollar[7]则发现成人VH、D和JH基因片段的使用是随机的,有较长的N区和高体突变存在。本研究首次发现,不同胎龄组新生儿IgH基因存在异型性:在早产儿,偏倚使用VH1和VH3,而在成熟儿(足月儿),则主要使用VH3 、VH5和 VH1。胎龄越小,NDN长度越短、体突变率越低,以及开放性阅读框越少。至于JH基因片段使用,在所有的新生儿,JH3的使用率均最高,JH2偶被使用,JH1未被使用;随着胎龄增加,JH3和JH6的使用率下降,JH4和JH5的使用率上升。在体突变的克隆中,大多数为置换突变A→G、C→G、T→C、G→T或C→T。综合Cuisinier、Choi和本研究的发现,可以推测:新生儿成熟度对IgH基因的重排产生明显影响,即随着新生儿的不断成熟,VH、D和JH基因片断的随机使用率,内在性体突变率、N区的长度,以及开放性阅读框等也增加,进而IgH基因多样性逐渐增加,表明人类新生儿和(或)胎儿体液免疫系统的发育也是一个渐进的发展过程。
, http://www.100md.com
基金项目:国家自然科学基金资助项目(39800175);广东省自然科学基金资助项目(980713)
参考文献
1,Tonegawa S. Somatic generation of antibody diversity. Nature, 1983, 302 :575-581.
2,Bhat NM, Bieber MM, Teng NN. Heavy chain variable gene usage by human B-1 lymphocytes and polyreactive autoantibodies. Hum Antibo, 1997, 8:146-150.
3,Ballow M, Cates KL, Rowe JC, et al. Development of the immune system in very low birth weight (less than 1 500g) premature infants: concentrations of plasma immunoglobulins and patterns of infections. Pediatr Res, 1986, 20:889-904.
, http://www.100md.com
4,Feeney AJ. Lack of N regions in fetal and neonatal mouse Ig V-D-J junctional sequences. J Exp Med, 1990, 172:1377-1390.
5,Sun J, Hayward C, Shinde R, et al. Antibody repertoire development in fetal and neonatal piglets. I. Four VH genes account for 80 percent of VH usage during 84 days of fetal life. J Immunol, 1998, 161:5070-5078.
6,Cuisinier AM, Guigou V, Boubli L, et al. Preferential expression of VH5 and VH6 immunoglobulin genes in early human B-cell ontogeny. Scand J Immunol, 1989, 30:493-497.
7,Stollar BD. The expressed heavy chain V gene repertorire of circulating B cells in normal adults. Ann N Y Acad Sci, 1995, 764:265-274.
(收稿日期:1999-07-08), 百拇医药