白细胞介素6在人骨髓瘤细胞系XG-7中的信号转导途径与生物学效应
作者:宋伦 沈倍奋
单位:军事医学科学院基础医学研究所 北京100850
关键词:白细胞介素6;多发性骨髓瘤;相对生物学效应
中华医学杂志001113
【摘要】 目的 研究白细胞介素6(IL-6)依赖型人骨髓瘤细胞系XG-7中IL-6信号转导途径活化与生物学效应之间的关系。方法 分别采用凝胶阻滞电泳和免疫沉淀方法检测参与IL-6信号传递功能的转录因子(STAT3、NF-IL-6)和蛋白激酶(JAK1、ERK)在XG-7细胞中的诱导激活状态并确定该细胞中的IL-6信号转导途径;进而采用细胞转染技术特异性上调或下调IL-6信号途径的诱导激活状态,并观察IL-6在XG-7细胞上生物学效应的改变。结果 IL-6可引发XG-7细胞中Ras/NF-IL-6信号转导途径活化,但没有出现Jak/STAT途径的诱导激活。Ras途径活化信号的上调与IL-6对XG-7细胞增殖促进作用的增强同步出现;而当Ras途径活化被抑制时,IL-6对XG-7细胞的生长促进作用明显减弱。结论 Ras/NF-IL-6信号转导途径介导了IL-6对XG-7细胞的生长维持和增殖促进作用。
, 百拇医药
Signal transduction and biological function of IL-6 in a human myeloma cell line-XG-7
SONG Lun SHEN Beifen
(Institute of Basic Medical Science, Academy of Military Medical Science, Beijing, 100850, China)
【Abstract】 Objective To investigate the relationship between IL-6 signal transduction and its biological function on a human IL-6-dependent myeloma cell line-XG-7. Methods Electrophoretic mobility shift (EMSA) and immunopeipitation were used respectively to detect the activation of transcription factors (STAT3 and NF-IL-6) and protein kinases (JAK1 and ERK) in the two IL-6 signal transduction pathways by IL-6 in XG-7 cells. Then the cells were transfected with the sense or anti-sense expression plasmids of the transcription factors, which could up- or down-regulate the activation of the signal transduction pathways specially, the changes of the biological function of IL-6 on XG-7 cells was shown by cell number counting and MTT. Results IL-6 can promote the proliferation of XG-7 cells and activate one of the IL-6 signal transduction pathway-Ras/NF-IL-6; but Jak/STAT signal transduction pathway was not activated at the same conditions. When the activation of Ras/NF-IL-6 pathway was up- or down-regulated, the growth effect of IL-6 on XG-7 cells was strongthened or weakened. Conclusions The proliferation of XG-7 cells in the presence of IL-6 is mediated by the activation of Ras/NF-IL-6 signal transduction pathway.
, 百拇医药
【Key words】 Interleukin-6; Multiple myeloma; Relative biological effectiveness
多发性骨髓瘤(multiple myeloma,MM)是一种晚期B细胞肿瘤,而IL-6是维持MM细胞生存和促进其增殖的一种最重要的细胞因子[1]。因此,研究IL-6在MM细胞中的信号转导途径与生物学效应之间的关系是揭示IL-6在MM中致病机制的有效方式。目前的研究结果所证实的IL-6信号转导途径主要有两条――Jak/STAT途径和Ras/NF-IL-6途径。在不同类型的IL-6靶细胞中,这两条信号途径的诱导激活状态可能存在差异,而且它们可能分别介导不同的IL-6生物学功能[2]。我们以一株IL-6生长依赖型MM细胞系――XG-7作为靶细胞,研究了与IL-6对该细胞的生长维持和增殖促进效应相对应的IL-6信号转导途径。
材料与方法
, 百拇医药
1.主要试剂:重组人IL-6由军事医学科学院生物工程研究所提供;α-32P-dATP购自北京福瑞公司;用于凝胶阻滞电泳法(EMSA)实验的寡核苷酸引物由本室合成(OPC柱纯化);抗ERK(胞外调节蛋白激酶)多克隆抗体和抗Jak1多克隆抗体购自Santa Cruz公司;抗磷酸酪氨酸抗体4G10为美国UBI公司产品,由美国NIH Stephen Shaw博士提供。
2.细胞系:人骨髓瘤细胞系XG-7[3]由苏州医学院张学光教授提供,用含有10%小牛血清的RPMI1640培养基传代培养,同时添加2 ng/ml的IL-6和5×10-5 mol/L的β-巯基乙醇。
3.IL-6对XG-7细胞生长的影响:用MTT方法检测,将传代培养的XG-7细胞于无血清1640培养基中饥饿4~6 h,然后以1×105个/ml的浓度接种入96孔板(每孔100 μl),并以每孔200 μl为终体积补加梯度稀释的IL-6和10% 1640培养基,72 h后加入MTT(5 mg/ml,每孔15 μl),代谢4~6 h后加入酸性SDS溶液溶解过夜并于570 nm处测定吸光值。
, 百拇医药
4.细胞核蛋白提取和凝胶阻滞电泳:参见文献[4]。凝胶阻滞电泳(EMSA)实验的DNA寡核苷酸探针为分别能够与转录因子STAT3结合的急性期反应成分和与NF-IL-6结合的NF-IL-6/激活转录因子。分别以“能够与活化的STAT3和NF-IL-6发生特异性结合”的DNA反应,急性期反应成分和NF-IL-6/激活转录因子作为探针,采用EMSA方法检测了IL-6刺激前后XG-7细胞中以上两种转录因子的诱导活化状态。
5.免疫沉淀:取XG-7细胞5×106~1×107个为一个样品,细胞预处理及刺激同核蛋白提取[4]。为了检测这两种蛋白激酶的诱导活化状态,我们首先采用抗ERK(或JAK1)抗体对IL-6刺激前后的细胞裂解物进行免疫沉淀,然后分别采用抗ERK(或JAK1)抗体和抗磷酸酪氨酸抗体4G10对免疫沉淀物进行识别。前者可检测出IL-6刺激前后目标蛋白ERK(或JAK1)在XG-7细胞中表达水平的变化;而后者可检测出这两种蛋白激酶的诱导活化情况。在刺激后的每一个细胞样品中加入0.25~0.5 ml的细胞裂解缓冲液(50 mmol/L Tris-Cl PH 7.5,150 mmol/L NaCl,1% NP40,0.5 mmol/L EDTA,0.5 mmol/L EGTA;临用前补加1 mmol/L Na3VO4、10 mmol/L NaF、2 mmol/L苯*****和10 μg/ml抑蛋白酶肽,抑制肽,亮抑制肽)冰浴作用30 min后,12 000 g离心20 min,弃沉淀,上清即为细胞裂解蛋白。在以上每份蛋白产物中加入抗ERK(或抗Jak1)抗体4 μg,4℃作用2 h,再加入prA-琼脂糖珠(美国Gibco公司)20 μl,4℃作用4 h或过夜。在进行电泳前将琼脂糖珠用洗涤缓冲液(除NP40浓度降至0.1%以外,其余成分同以上裂解缓冲液)和PBS溶液各洗3遍,然后走10% SDS-PAGE电泳,按常规方法转印,并分别用抗ERK(或抗Jak1)和抗磷酸酪氨酸抗体(a-P-Tyr)4G10进行识别,用AP标记的二抗及其底物5-溴-4-氯-3-吲哚磷酸盐、氮蓝四唑显色。
, http://www.100md.com
6.表达质粒转染XG-7细胞:采用美国Gibco公司Lipofactamine转染试剂并按其产品说明书方法进行。
结果
1.IL-6对XG-7细胞的生长维持和增殖促进作用:XG-7细胞为1994年建系的人MM细胞系[3],在没有IL-6存在时,XG-7细胞基本不能存活;随着IL-6刺激浓度的增加,XG-7细胞的增殖效应逐渐增强(图1)。因此XG-7细胞属于IL-6生长依赖型人MM细胞系。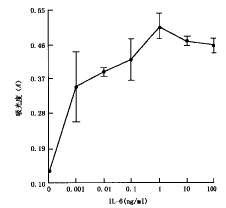
图1 IL-6对XG-7细胞生长的影响
2.IL-6诱导XG-7细胞中转录因子NF-IL-6活化:在两条IL-6信号转导途径中,转录因子STAT3和NF-IL-6只有被IL-6诱导激活后才能各自形成同二聚体,进而与相应的DNA反应成分结合并诱导靶基因表达[2]。
, http://www.100md.com
不同剂量IL-6刺激XG-7细胞后没有出现STAT3活化信号;而NF-IL-6在IL-6刺激前即处于组成性激活状态,IL-6刺激后其活化信号明显增强(图2A)。同一有效IL-6剂量(10 ng/ml)在刺激不同时间点的电泳结果(图2B)也显示出,NF-IL-6可在IL-6刺激15 min之内出现一过性激活信号,30 min后活化信号减弱,60 min后又恢复至未刺激前水平;而STAT3在各个时间点没有相应的活化信号出现。
3.IL-6诱导XG-7细胞中蛋白激酶ERK活化:在两条IL-6信号转导途径的级联活化反应中,催化转录因子NF-IL-6和STAT3活化的蛋白激酶分别是ERK(MAPK)和Jak。抗ERK抗体的识别可检测出IL-6刺激前后XG-7细胞中表达量无明显改变的目标蛋白ERK1和ERK2;而抗磷酸酪氨酸抗体4G10可识别出IL-6刺激前组成性激活的ERK(微弱)以及IL-6刺激后活化信号显著增强的ERK;同样条件下,没有检测到Jak1的表达及诱导活化信号(图3)。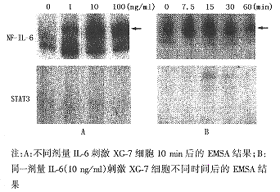
, 百拇医药
注:A:不同剂量IL-6刺激XG-7细胞10 min后的EMSA结果;B:同一剂量IL-6(10 ng/ml)刺激XG-7细胞不同时间后的EMSA结果
图2 STAT3和NF-IL-6在XG-7细胞中的诱导激活情况
图3 蛋白激酶Jak1和ERK在XG-7细胞中的诱导活化情况
4.NF-IL-6正义及反义表达质粒对XG-7细胞增殖和IL-6信号转导功能的影响:将NF-IL-6 cDNA分别正向和反向插入到真核表达载体pCDNA3和pSVL中构建了NF-IL-6正义表达质粒pCD-NF-IL-6和反义表达质粒pAT-NF-IL-6。将这两个表达质粒与它们分别对应的空载体对照转染XG-7细胞,结果转染pAT-NF-IL-6的XG-7细胞中NF-IL-6诱导活化信号明显减弱;而pCD-NF-IL-6转染后的XG-7细胞NF-IL-6的活化信号显著增强(图4),这说明这两个表达质粒可分别下调和上调Ras途径的诱导激活。对转染后的细胞计数,结果显示,转染pAT-NF-IL-6的XG-7细胞的生长被明显抑制;而pCD-NF-IL-6的导入使XG-7细胞获得了比对照组更强的生长能力(图5)。说明当Ras途径活化被上调或下调时,XG-7细胞的生长分别得以增强和减弱。pAT-NF-IL-6转染后的XG-7细胞对IL-6的反应敏感性显著下降(图6)。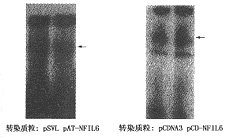
, 百拇医药
图4 pAT-NF-IL-6和pCD-NF-IL-6转染XG-7细胞后的EMSA检测结果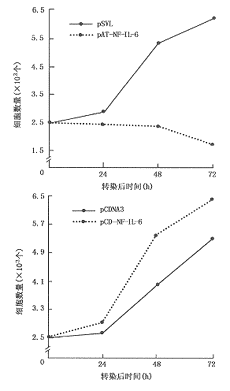
图5 pAT-NF-IL-6和pCD-NF-IL-6转染后XG-7细胞生长情况的改变
讨论
骨髓瘤细胞的生存和增殖与IL-6关系密切。在MM病程发展初期(即非活化期),恶变型成熟浆细胞迁移至骨髓内,并依赖骨髓基质细胞分泌的IL-6使之得以存活[1]。因此,本研究所选用的“IL-6生长依赖型MM细胞系――XG-7”可以基本反映MM病程发展初期体内瘤细胞的生存状态。经过我们的研究发现,XG-7细胞在生存过程中存在有Ras/NF-IL-6信号转导途径的组成性激活;在IL-6刺激后,Ras途径的活化信号进一步增强,并且信号途径活化状态的改变与IL-6对该细胞增殖促进作用的改变同步出现;因此,Ras途径的活化介导了IL-6对XG-7细胞的生存维持和增殖促进效应。但我们注意到,在同样的IL-6刺激作用下,XG-7细胞中没有出现JAK/STAT途径的诱导激活。此结果提示,可能在MM发育早期,IL-6的刺激作用主要引发了Ras/NF-IL-6信号转导途径活化,并介导了此阶段IL-6在MM细胞上的生物学效应;而JAK/STAT途径则不参与处于早期发育阶段的MM细胞中的IL-6信号转导功能。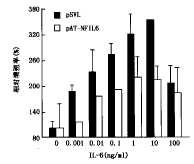
, http://www.100md.com
图6 pAT-NF-IL-6转染后XG-7细胞IL-6增殖反应性的改变
参考文献
1,Hallek M, Bergsagel PL, Anderson KC. Multiple myeloma: increasing evidence for a multistep transformation process. Blood, 1998,91:3-21.
2,宋伦,沈倍奋.IL-6信号转导的分子机制.《国外医学》免疫学分册,1999,22:79-83.
3,Zhang XG, Gu ZJ, Lu ZY, et al. Reproducible obtaining of human myeloma cell lines as a model for tumer stem cell study in human multiple myeloma. Blood, 1994,83:3654-3660.
4,宋伦,黎燕,沈倍奋.280位His残基对sIL-6R介导的IL-6信号转导功能的影响.中国实验临床免疫学杂志,1999,5:29-32.
(收稿日期:2000-03-08), http://www.100md.com
单位:军事医学科学院基础医学研究所 北京100850
关键词:白细胞介素6;多发性骨髓瘤;相对生物学效应
中华医学杂志001113
【摘要】 目的 研究白细胞介素6(IL-6)依赖型人骨髓瘤细胞系XG-7中IL-6信号转导途径活化与生物学效应之间的关系。方法 分别采用凝胶阻滞电泳和免疫沉淀方法检测参与IL-6信号传递功能的转录因子(STAT3、NF-IL-6)和蛋白激酶(JAK1、ERK)在XG-7细胞中的诱导激活状态并确定该细胞中的IL-6信号转导途径;进而采用细胞转染技术特异性上调或下调IL-6信号途径的诱导激活状态,并观察IL-6在XG-7细胞上生物学效应的改变。结果 IL-6可引发XG-7细胞中Ras/NF-IL-6信号转导途径活化,但没有出现Jak/STAT途径的诱导激活。Ras途径活化信号的上调与IL-6对XG-7细胞增殖促进作用的增强同步出现;而当Ras途径活化被抑制时,IL-6对XG-7细胞的生长促进作用明显减弱。结论 Ras/NF-IL-6信号转导途径介导了IL-6对XG-7细胞的生长维持和增殖促进作用。
, 百拇医药
Signal transduction and biological function of IL-6 in a human myeloma cell line-XG-7
SONG Lun SHEN Beifen
(Institute of Basic Medical Science, Academy of Military Medical Science, Beijing, 100850, China)
【Abstract】 Objective To investigate the relationship between IL-6 signal transduction and its biological function on a human IL-6-dependent myeloma cell line-XG-7. Methods Electrophoretic mobility shift (EMSA) and immunopeipitation were used respectively to detect the activation of transcription factors (STAT3 and NF-IL-6) and protein kinases (JAK1 and ERK) in the two IL-6 signal transduction pathways by IL-6 in XG-7 cells. Then the cells were transfected with the sense or anti-sense expression plasmids of the transcription factors, which could up- or down-regulate the activation of the signal transduction pathways specially, the changes of the biological function of IL-6 on XG-7 cells was shown by cell number counting and MTT. Results IL-6 can promote the proliferation of XG-7 cells and activate one of the IL-6 signal transduction pathway-Ras/NF-IL-6; but Jak/STAT signal transduction pathway was not activated at the same conditions. When the activation of Ras/NF-IL-6 pathway was up- or down-regulated, the growth effect of IL-6 on XG-7 cells was strongthened or weakened. Conclusions The proliferation of XG-7 cells in the presence of IL-6 is mediated by the activation of Ras/NF-IL-6 signal transduction pathway.
, 百拇医药
【Key words】 Interleukin-6; Multiple myeloma; Relative biological effectiveness
多发性骨髓瘤(multiple myeloma,MM)是一种晚期B细胞肿瘤,而IL-6是维持MM细胞生存和促进其增殖的一种最重要的细胞因子[1]。因此,研究IL-6在MM细胞中的信号转导途径与生物学效应之间的关系是揭示IL-6在MM中致病机制的有效方式。目前的研究结果所证实的IL-6信号转导途径主要有两条――Jak/STAT途径和Ras/NF-IL-6途径。在不同类型的IL-6靶细胞中,这两条信号途径的诱导激活状态可能存在差异,而且它们可能分别介导不同的IL-6生物学功能[2]。我们以一株IL-6生长依赖型MM细胞系――XG-7作为靶细胞,研究了与IL-6对该细胞的生长维持和增殖促进效应相对应的IL-6信号转导途径。
材料与方法
, 百拇医药
1.主要试剂:重组人IL-6由军事医学科学院生物工程研究所提供;α-32P-dATP购自北京福瑞公司;用于凝胶阻滞电泳法(EMSA)实验的寡核苷酸引物由本室合成(OPC柱纯化);抗ERK(胞外调节蛋白激酶)多克隆抗体和抗Jak1多克隆抗体购自Santa Cruz公司;抗磷酸酪氨酸抗体4G10为美国UBI公司产品,由美国NIH Stephen Shaw博士提供。
2.细胞系:人骨髓瘤细胞系XG-7[3]由苏州医学院张学光教授提供,用含有10%小牛血清的RPMI1640培养基传代培养,同时添加2 ng/ml的IL-6和5×10-5 mol/L的β-巯基乙醇。
3.IL-6对XG-7细胞生长的影响:用MTT方法检测,将传代培养的XG-7细胞于无血清1640培养基中饥饿4~6 h,然后以1×105个/ml的浓度接种入96孔板(每孔100 μl),并以每孔200 μl为终体积补加梯度稀释的IL-6和10% 1640培养基,72 h后加入MTT(5 mg/ml,每孔15 μl),代谢4~6 h后加入酸性SDS溶液溶解过夜并于570 nm处测定吸光值。
, 百拇医药
4.细胞核蛋白提取和凝胶阻滞电泳:参见文献[4]。凝胶阻滞电泳(EMSA)实验的DNA寡核苷酸探针为分别能够与转录因子STAT3结合的急性期反应成分和与NF-IL-6结合的NF-IL-6/激活转录因子。分别以“能够与活化的STAT3和NF-IL-6发生特异性结合”的DNA反应,急性期反应成分和NF-IL-6/激活转录因子作为探针,采用EMSA方法检测了IL-6刺激前后XG-7细胞中以上两种转录因子的诱导活化状态。
5.免疫沉淀:取XG-7细胞5×106~1×107个为一个样品,细胞预处理及刺激同核蛋白提取[4]。为了检测这两种蛋白激酶的诱导活化状态,我们首先采用抗ERK(或JAK1)抗体对IL-6刺激前后的细胞裂解物进行免疫沉淀,然后分别采用抗ERK(或JAK1)抗体和抗磷酸酪氨酸抗体4G10对免疫沉淀物进行识别。前者可检测出IL-6刺激前后目标蛋白ERK(或JAK1)在XG-7细胞中表达水平的变化;而后者可检测出这两种蛋白激酶的诱导活化情况。在刺激后的每一个细胞样品中加入0.25~0.5 ml的细胞裂解缓冲液(50 mmol/L Tris-Cl PH 7.5,150 mmol/L NaCl,1% NP40,0.5 mmol/L EDTA,0.5 mmol/L EGTA;临用前补加1 mmol/L Na3VO4、10 mmol/L NaF、2 mmol/L苯*****和10 μg/ml抑蛋白酶肽,抑制肽,亮抑制肽)冰浴作用30 min后,12 000 g离心20 min,弃沉淀,上清即为细胞裂解蛋白。在以上每份蛋白产物中加入抗ERK(或抗Jak1)抗体4 μg,4℃作用2 h,再加入prA-琼脂糖珠(美国Gibco公司)20 μl,4℃作用4 h或过夜。在进行电泳前将琼脂糖珠用洗涤缓冲液(除NP40浓度降至0.1%以外,其余成分同以上裂解缓冲液)和PBS溶液各洗3遍,然后走10% SDS-PAGE电泳,按常规方法转印,并分别用抗ERK(或抗Jak1)和抗磷酸酪氨酸抗体(a-P-Tyr)4G10进行识别,用AP标记的二抗及其底物5-溴-4-氯-3-吲哚磷酸盐、氮蓝四唑显色。
, http://www.100md.com
6.表达质粒转染XG-7细胞:采用美国Gibco公司Lipofactamine转染试剂并按其产品说明书方法进行。
结果
1.IL-6对XG-7细胞的生长维持和增殖促进作用:XG-7细胞为1994年建系的人MM细胞系[3],在没有IL-6存在时,XG-7细胞基本不能存活;随着IL-6刺激浓度的增加,XG-7细胞的增殖效应逐渐增强(图1)。因此XG-7细胞属于IL-6生长依赖型人MM细胞系。
图1 IL-6对XG-7细胞生长的影响
2.IL-6诱导XG-7细胞中转录因子NF-IL-6活化:在两条IL-6信号转导途径中,转录因子STAT3和NF-IL-6只有被IL-6诱导激活后才能各自形成同二聚体,进而与相应的DNA反应成分结合并诱导靶基因表达[2]。
, http://www.100md.com
不同剂量IL-6刺激XG-7细胞后没有出现STAT3活化信号;而NF-IL-6在IL-6刺激前即处于组成性激活状态,IL-6刺激后其活化信号明显增强(图2A)。同一有效IL-6剂量(10 ng/ml)在刺激不同时间点的电泳结果(图2B)也显示出,NF-IL-6可在IL-6刺激15 min之内出现一过性激活信号,30 min后活化信号减弱,60 min后又恢复至未刺激前水平;而STAT3在各个时间点没有相应的活化信号出现。
3.IL-6诱导XG-7细胞中蛋白激酶ERK活化:在两条IL-6信号转导途径的级联活化反应中,催化转录因子NF-IL-6和STAT3活化的蛋白激酶分别是ERK(MAPK)和Jak。抗ERK抗体的识别可检测出IL-6刺激前后XG-7细胞中表达量无明显改变的目标蛋白ERK1和ERK2;而抗磷酸酪氨酸抗体4G10可识别出IL-6刺激前组成性激活的ERK(微弱)以及IL-6刺激后活化信号显著增强的ERK;同样条件下,没有检测到Jak1的表达及诱导活化信号(图3)。
, 百拇医药
注:A:不同剂量IL-6刺激XG-7细胞10 min后的EMSA结果;B:同一剂量IL-6(10 ng/ml)刺激XG-7细胞不同时间后的EMSA结果
图2 STAT3和NF-IL-6在XG-7细胞中的诱导激活情况
图3 蛋白激酶Jak1和ERK在XG-7细胞中的诱导活化情况
4.NF-IL-6正义及反义表达质粒对XG-7细胞增殖和IL-6信号转导功能的影响:将NF-IL-6 cDNA分别正向和反向插入到真核表达载体pCDNA3和pSVL中构建了NF-IL-6正义表达质粒pCD-NF-IL-6和反义表达质粒pAT-NF-IL-6。将这两个表达质粒与它们分别对应的空载体对照转染XG-7细胞,结果转染pAT-NF-IL-6的XG-7细胞中NF-IL-6诱导活化信号明显减弱;而pCD-NF-IL-6转染后的XG-7细胞NF-IL-6的活化信号显著增强(图4),这说明这两个表达质粒可分别下调和上调Ras途径的诱导激活。对转染后的细胞计数,结果显示,转染pAT-NF-IL-6的XG-7细胞的生长被明显抑制;而pCD-NF-IL-6的导入使XG-7细胞获得了比对照组更强的生长能力(图5)。说明当Ras途径活化被上调或下调时,XG-7细胞的生长分别得以增强和减弱。pAT-NF-IL-6转染后的XG-7细胞对IL-6的反应敏感性显著下降(图6)。
, 百拇医药
图4 pAT-NF-IL-6和pCD-NF-IL-6转染XG-7细胞后的EMSA检测结果
图5 pAT-NF-IL-6和pCD-NF-IL-6转染后XG-7细胞生长情况的改变
讨论
骨髓瘤细胞的生存和增殖与IL-6关系密切。在MM病程发展初期(即非活化期),恶变型成熟浆细胞迁移至骨髓内,并依赖骨髓基质细胞分泌的IL-6使之得以存活[1]。因此,本研究所选用的“IL-6生长依赖型MM细胞系――XG-7”可以基本反映MM病程发展初期体内瘤细胞的生存状态。经过我们的研究发现,XG-7细胞在生存过程中存在有Ras/NF-IL-6信号转导途径的组成性激活;在IL-6刺激后,Ras途径的活化信号进一步增强,并且信号途径活化状态的改变与IL-6对该细胞增殖促进作用的改变同步出现;因此,Ras途径的活化介导了IL-6对XG-7细胞的生存维持和增殖促进效应。但我们注意到,在同样的IL-6刺激作用下,XG-7细胞中没有出现JAK/STAT途径的诱导激活。此结果提示,可能在MM发育早期,IL-6的刺激作用主要引发了Ras/NF-IL-6信号转导途径活化,并介导了此阶段IL-6在MM细胞上的生物学效应;而JAK/STAT途径则不参与处于早期发育阶段的MM细胞中的IL-6信号转导功能。
, http://www.100md.com
图6 pAT-NF-IL-6转染后XG-7细胞IL-6增殖反应性的改变
参考文献
1,Hallek M, Bergsagel PL, Anderson KC. Multiple myeloma: increasing evidence for a multistep transformation process. Blood, 1998,91:3-21.
2,宋伦,沈倍奋.IL-6信号转导的分子机制.《国外医学》免疫学分册,1999,22:79-83.
3,Zhang XG, Gu ZJ, Lu ZY, et al. Reproducible obtaining of human myeloma cell lines as a model for tumer stem cell study in human multiple myeloma. Blood, 1994,83:3654-3660.
4,宋伦,黎燕,沈倍奋.280位His残基对sIL-6R介导的IL-6信号转导功能的影响.中国实验临床免疫学杂志,1999,5:29-32.
(收稿日期:2000-03-08), http://www.100md.com