调整Th1/Th2平衡可用于肾脏疾病的治疗吗?
作者:姚小丹 黎磊石
单位:南京军区南京总医院 解放军肾脏病研究所(南京,210002)
关键词:Th1/Th2;肾脏病;治疗
肾脏病与透析肾移植杂志990329
自从1986年Mosmman和Coffman依据分泌细胞因子的差别将小鼠CD4+T细胞进一步分成Th1(主要产生IL-2及IFN-γ)及Th2(主要产生IL-4及ΙL-5)细胞亚群以来[1],Th1和Th2细胞效应及其平衡,在实验动物模型及人类感染和自身免疫性疾病中得到了深入研究。研究肾脏疾病中Th1与Th2平衡的意义,不仅在于更深入地理解疾病免疫发病机制,更重要的在于探索肾脏疾病治疗新途径[2]。那么,调整Th1/Th2平衡能否用于肾脏疾病的临床治疗呢?
, http://www.100md.com
1 Th1/Th2细胞效应的特征及其平衡
Th1细胞效应所产生的细胞因子除了IL-2与IFN-γ外,还包括TNF-β和IL-17(后者也是一种促炎症因子,能增加IL-6,GM-CSF及PGE2的产生)。Th2细胞效应所产生的细胞因子除了IL-4及IL-5外,还包括IL-6,IL-10及IL-13。Th1细胞效应主要是诱导巨噬细胞及细胞毒性T淋巴细胞活化,B细胞所产生的IgG转向(swith)易于补体活化及发挥调理作用的IgG亚类(小鼠的IgG2a和IgG3,人类的IgG1和IgG3),因此Th1细胞效应主要促进细胞内致病原的清除及器官特异性自身免疫反应(如迟发型变态反应DTH);而Th2细胞效应所产生的细胞因子(IL-4,IL-5,IL-10)参与了过敏性变态反应及寄生虫感染性疾病,促进B细胞产生IgE及IgG1(小鼠)或IgG4(人类)[3]。Th1和Th2的前体细胞(Tho)本身也是由原始T细胞活化而来,本身又能产生IL-4、IL-2和IFN-γ,原始T细胞活化后能产生IL-2。最近,Th3细胞也为人们所认识,它所产生的主要细胞因子为TGF-β[4]。
, http://www.100md.com
Th1/Th2细胞效应的特征之一是自身促进作用,即各亚群Th细胞因子诱导Th0细胞向各自的细胞亚群分化并促进各自Th细胞亚群活化。IFN-γ诱导Th0细胞向Th1细胞亚群分化,IL-4诱导Th0细胞向Th2细胞亚群分化[5]。
Th1/Th2细胞效应的特征之二是相互抑制作用,即各类Th细胞因子对另一类细胞亚群的活化及细胞因子的产生具有抑制作用。IL-4及IL-10都是IFN-γ的强力抑制剂,而IFN-γ亦反过来能抑制IL-10的产生[6]。
Th1/Th2细胞效应在通常情况下保持着相对的平衡(附图)[7]。这一平衡的维持是相当错综复杂的,就是在疾病状态下,Th1/Th2功能的消长也不完全取决于Th细胞本身的功能及所产生的细胞因子。对这一平衡的研究表明,虽然启动Th1/Th2细胞效应抗原各不相同,但是,通过改变细胞因子环境及采用丝裂原刺激,在体内或体外即能改变Th0细胞的分化方向。在丝裂原刺激下,胎儿脐血T细胞如培养在IL-12及抗IL-4环境中则向Th1细胞分化;而培养在IL-4及抗IL-12的环境中则向Th2细胞分化,这提示调整Th1/Th2细胞效应平衡是可能的,而且可以绕过特异性抗原来调整这一平衡[8]。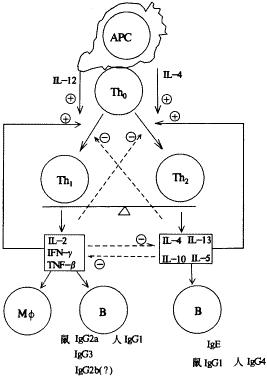
, http://www.100md.com
附图 Th1/Th2细胞效应与细胞因子间的平衡及其相互作用
实线为促进或正反馈作用,虚线为抑制或负反馈作用
2 Th1/Th2细胞效应的调控途径
目前人们已认识到Th1/Th2细胞效应主要受以下环节的影响。
2.1 基因背景 Th1/Th2细胞效应的平衡受到诸多基因的影响。比如,在Th1细胞效应中,早期巨噬细胞分泌的IL-12是重要的决定因子,若在免疫反应启动初期缺乏抗原呈递细胞(APC)分泌的IL-12的作用,则T细胞可能因表达的IL-12受体中缺失β2亚单位而不再对IL-12发生进一步的反应。这种不反应性与11号染色体的一个区域(在人类为5p31区)有关联。实验性自身免疫性脑脊髓炎和非肥胖1型糖尿病的易感性也与此区域有关联。
2.2 抗原呈递 抗原的呈递(包括呈递的类型、剂量和途径)与Th1/Th2细胞效应有重要关系。采用极低剂量的抗原于粘膜或腹腔内刺激,可以诱导Th2细胞效应以产生抗炎细胞因子。如在类风显性关节炎(RA)患者采用小剂量胶原口服,可能诱导免疫耐受而治疗RA[9,11]。
, http://www.100md.com
2.3 细胞因子环境 如上所述,细胞因子环境对于两类细胞的分化具有肯定的影响,目前可以通过增强内源性细胞因子的表达及提供外源性细胞因子,或(和)采用相应细胞因子的拮抗因子,以改变细胞因子环境而调整Th1/Th2平衡。
2.4 细胞活化协同刺激分子 抗原呈递细胞表面表达的协同刺激分子B7-1(CD80)及B7-2(CD86)是两类Th细胞分化过程中的第二信使,两信使分子的T细胞表面配体为CTLA-4及CD28。小鼠体内研究证实,激活或阻断B7-1途径,可促进或抑制Th1细胞效应,而激活或阻断B7-2途径,可促进或抑制Th2细胞效应。
2.5 细胞活化信号通路 IL-12的作用需要经过Stat4基因的活化与表达,小鼠Stat4基因的不活化可导致IL-12产生障碍,最后引起Th1细胞效应障碍,而Stat6基因敲除小鼠IL-4的产生及Th2细胞效应亦出现明显障碍。IL-12受体中β2亚单位的表达是Th1细胞效应中对IL-12反应性的关键,而调节IL-12R β2可以控制以Th1细胞效应为主导的自身免疫性疾病的发生。
, 百拇医药
3 Th1及Th2平衡与肾脏疾病及其它自身免疫性疾病
当前判断疾病免疫发病机制中Th1或Th2细胞效应为主导的方法,主要是依据炎症组织局部细胞因子的表达,也可根据外周血清中抗体反应,如IgE的水平及IgG亚类的变化进行判断。在动物模型的研究中,根据干扰Th1或Th2细胞效应的各种环节后Th1或Th2细胞效应的变化来进行判断(见附表)。
附表 与Th1或Th2平衡相关的人类感染疾病及自免疫性疾病[9~11] Th1主导
Th1/Th2
Th2主导
人类肾脏病
伴新月体形成的弥漫增生性肾炎
, 百拇医药 IgA肾炎
膜性肾病
抗GBM疾病
狼疮性肾炎
微小病变
寡免疫复合物型新月体肾炎
Ⅴ型狼疮性肾炎
膜增殖性肾炎
类风湿性关节炎相关性肾炎
IFN-γ治疗后伴新月体形成的狼疮性肾炎
IL-2治疗后伴新月体形成的IgA肾炎
肾炎动物模型
, 百拇医药
异质期抗GBM肾炎的C57BL/6小鼠
抗GBM肾炎BALB/c小鼠(较少有新月体形成)
巨噬细胞依赖性抗GBM肾炎的Wistar-Kyoto大鼠
抗GBM肾炎的SD大鼠
狼疮性肾炎鼠模型
氯化汞诱发的肾小球肾炎鼠或兔模型
慢性GVH疾病
伴有膜性肾病改变的慢性HVG疾病
Hymann肾炎
其它疾病或情形
, 百拇医药 多发性硬化
哮喘
自身免疫性甲状腺炎
过敏性皮炎
重症眼肌无力
季节性结膜炎
类风湿性关节炎
正常妊娠免疫耐受
Lyme关节炎
特发性嗜酸性细胞增多症
反应性关节炎
SLE
, 百拇医药 接触性皮炎
硬皮病
1型糖尿病
慢性GVH病
结节性红斑
Omenn's Syndrome
习惯性流产
GVH:宿主抗移植物,HVG:移植抗宿主
4 肾脏疾病中针对Th1及Th2平衡的治疗措施
在Th1主导的实验性自身免疫性脑脊髓炎,自身免疫性甲状腺炎,1型糖尿病非肥胖小鼠模型及实验性结肠炎动物模型中,采用外源性IL-4或(和)IL-10治疗,能够选择性地抑制Th1细胞效应而减轻疾病的严重程度。这类研究的结果,为调整Th1/Th2细胞效应平衡提供了实验基础[9]。
, 百拇医药
4.1 直接针对Th1或Th2细胞功能与分化的措施 由于目前对于Th1及Th2细胞表面标记分子(如CD27及CD30)研究中存在有较多争议,因而直接作用于细胞表面分子的单克隆抗体的措施尚无任何报道。但是,T细胞IL-12受体的β2亚单位的表达与Th1细胞效应有比较密切的关系,而GATA-3与Th2细胞效应有比较密切的关系,而在各类肾脏疾病中观察这两种分子的表达有利于判断Th1与Th2平衡,并为相应的干预措施提供监测依据。另外,体外应用细胞因子激活的Th1或Th2细胞,然后转输注体内,也可能成为直接干预Th1或Th2平衡的措施之一。这两种可能的治疗措施尚未在动物模型上应用。
4.2 针对细胞因子平衡的措施
4.2.1 直接针对细胞因子 针对细胞因子的措施包括单独或联合投用外源性细胞因子(如IL-4,IL-6及IL-10,TGF-β),或其拮抗剂(如抗IL-10及抗IL-4的中和性抗体),或采用抗炎因子基因疗法。其中最有潜力的是Th2细胞因子系列中的IL-4和IL-10,而IL-6及TGF-β治疗作用相对不确定而潜力较小。基因疗法尚在初步的探讨中。
, http://www.100md.com
Tipping等[12]在异质期抗GBM肾炎的C57BL/6小鼠模型上证实,联合应用IL-4及IL-10可减少新月体的形成及保护肾功能,其中IL-10具有独立的保护功能,而IL-4不具有独立的保护作用。这一保护作用很可能不完全是Th1向Th2转化的结果,而是IL-4及IL-10本身的抗炎作用。上述假设在巨噬细胞依赖性抗GBM肾炎的Wistar-Kyoto大鼠模型上得以验证,肾小球中巨噬细胞活化标记在IL-10治疗后也相应减少。
Tam等[13]在进展型抗GBM非新月体肾炎SD大鼠模型上证实,在抗原刺激之前采用IL-4治疗可以大幅度减少蛋白尿及肾脏病理损伤。这种作用与局部Ⅱ型IL-1结合性受体表达增多而结合IL-1,致使后者对浸润巨噬细胞的刺激减弱有关。
在Fouqueray[14]的研究中,应用人类IL-10基因转染(通过转染肾系膜细胞或全身骨骼肌细胞)至同质期抗GBM大鼠模型后,蛋白尿排泄减少。
, 百拇医药
但Chadban等[15]的研究指出,自注射抗GBM抗体后,每天投用重组鼠IL-10治疗SD大鼠模型并不能减少新月体的形成,相反,动物还会产生抗鼠IL-10的免疫反应,后者则可能加剧肾脏的损伤。
在多数抗GBM疾病的模型中,IL-4治疗的确具有肯定的抗炎作用,但其抗炎作用并不完全是通过作用Th1与Th2平衡发生的,还可能与抗炎脂质分子的产生有关。不过现在对于IL-4治疗的副作用知之甚少,IL-4能加强VCAM-1表达,从而加强T细胞及嗜酸性粒细胞与内皮细胞的粘附,IL-4还能刺激细胞外基质的产生,增加纤维母细胞产生 Ⅰ 型、Ⅲ 型胶原及纤维结合蛋白,这样可能引起T细胞在炎症局部的聚集和炎症局部组织的硬化。在Heymann肾炎中,增强IL-4 mRNA表达与肾小球中T细胞浸润无肯定的关系。
服用IL-10(而不是IL-4)对于大鼠抗GBM肾炎有保护作用,IL-10的保护作用更大程度上归因于它对Th1及Th2平衡的调整效应。因为在IL-10治疗后,从属Th1系列的免疫球蛋白亚类(IgG2a和IgG3)产生减少,而从属Th2系列的免疫球蛋白亚类(IgG1)的产生变化不明显。所以IL-10是目前所知的潜力最大的治疗制剂,已用于RA的Ⅱ期临床治疗[11]。不过,长期应用IL-10可引起Th2主导的肾小球肾炎的发作,如动物模型(NZB/WF1小鼠)治疗后发生狼疮样肾炎。而采用抗IL-10可延迟这类肾炎的发生。TGF-β与IL-10的抗炎作用相似。在实验性系膜增殖性肾炎模型中[16],TGF-β1的基因治疗可以在肾小球组织中表达足够活性的TGF-β,从而抑制肾小球细胞的增殖反应,这一作用可能与巨噬细胞分泌的炎症因子减少有关。
, http://www.100md.com
多数IgA肾炎患者存在Th1及Th2细胞效应的证据,但高血清IgA(特别是大分子IgA)患者的Th2效应占主导。IL-4不仅参与了免疫球蛋白的转向作用,而且还发挥了旁分泌与自分泌的作用,参与了IgA的沉积过程。而在伴新月体形成的IgA肾炎中,IgG1、IgG3沉积突出,且IFN-γ表达增强(Th1效应)。但目前对IgA肾炎尚无干扰尝试的报道。
在SLE中,单纯Th1或Th2效应的证据是难以确立的,但对不同类型狼疮性肾炎,Th1或Th2效应的主次仍有差别。在动物模型中,无论是单独增强Th1还是Th2细胞因子的作用,都可能加重病情,服用IFN-γ或IL-12也不能减少自身抗体的产生,但无论是阻断Th1还是Th2细胞因子,都可以减轻病情[10]。
在氯化汞诱导的肾小球肾炎动物模型中,Th2效应占主导地位。应用IFN-γ治疗这种肾炎小鼠虽然能减少IgE的产生,但不能减少自身抗体的产生及阻止肾炎的形成。
, http://www.100md.com
肾脏固有细胞虽然也能产生部分Th1与Th2系列的细胞因子,但是现在尚没有证据表明致肾炎性免疫反应的发生与肾脏固有细胞产生的细胞因子有关。在炎症过程中,肾脏固有细胞与免疫细胞之间的确有相互作用,但目前更多的人认为是全身性免疫反应决定了Th1与Th2效应。
4.2.2 诱导体内保护性细胞因子 尽管采用外源性针对细胞因子的措施可以取得肯定的作用,但是对慢性自身免疫性疾病来说,长期体外服用细胞因子是不现实的,而且抗原长期刺激的体外研究已表明维持长期Th1与Th2间的转化是不可能的。另外,疾病早期与后期的Th1/Th2平衡可能不尽相似。IL-12在反复发作的感染性疾病及自身免疫性疾病的复发过程中起着重要的作用,而应用抗IL-12在Th1主导的疾病发生早期或复发前,可能阻止其发作或复发,而疾病发生后应用抗IL-12则可能加重Th1主导的疾病的程度。
4.3 诱导旁观抑制效应 激活的Th2/Th3细胞对于Th1细胞依赖的炎症反应具有保护应用,这一作用的发生主要是通过抗原刺激下的IL-4,IL-10及TGF-β而发挥的。如在粘膜表面采用一定剂量的抗原摄入可诱导Th3细胞的产生,这一作用不依赖于IL-4,而依赖于TGF-β,而采用修饰抗原刺激则出现依赖于IL-4的途径。不过,这样激活的T细胞主要位于粘膜局部,而使这类细胞迁移至炎症局部还有没能解决的困难。有人发现在早期的抗原接触过程中,应用非耗竭性抗CD4多克隆抗体可以使Th1细胞效应转化为产生IL-4的Th2细胞效应[17]。
, 百拇医药
总之,对于Th1/Th2平衡与肾脏疾关系的研究正在不断深入,而对于这一平衡的治疗措施,尚处于动物实验的初级阶段。就是动物模型上,这些措施多数仅在疾病发作的早期有治疗效应。应用于临床肾脏疾病的治疗仍有不少尚未克服的问题。不过,这一领域的研究进展日新月异,部分细胞因子生物制剂已用于RA的Ⅱ期临床研究。调整Th1/Th2效应平衡在肾脏病治疗中的应用指日可待。
参考文献
1 Mosmann TR,Cherwinski H,Bond MW et al.Two type of murine helper T cell clone.I.Definition according to profies of activities and secreted proteins.J Imunol,1986,136:2348
2 Baud L,Fouqueray B,Bellocq A.Switching off renal inflammation by anti-inflammatory mediators:the facts,the promise and the hope.Kidney Int,1998,53:1118
, 百拇医药
3 Romagnani S.Human Th1 and Th2 subsets:doubt no more.Immunol Today,1991,12:256
4 Abbas AK,Murphy KM,Sher A.Functional diversity of helper T lymphocytes.Nature,1996,383:787
5 Daynes RA,Araneo BA,Dowell TA et al.Regulation of murine lymphokine production in vivo.Ⅲ.The lymphoid tissue microenvironment exerts regulatory influences over T helper cell function.J Exp Med,1990,171:979
6 Chomarat P,Issoan MC,Banchereau J et al.Interferon γ inhibits interleukin-10 production by monocytes.J Exp Med,1993,177;523
, http://www.100md.com
7 刘俊铎,刘志红.Th1/Th2细胞与自身免疫性疾病.肾脏病与透析肾移植杂志,1997,6(6):555
8 Sszabo SJ,Dighe AS,Gubler U et al.Regulation of the interleukin-12R β 2subunit expression in developing T helper 1 and Th2 cells.J Exp Med,1997,185:817
9 Miossec P,van den Berg W.Th1/Th2 cytokine balance in arthritis.Arthritis Rheum,1997,40(12):2105
10 Holdsworth SR,Kitching R,Tipping PG.Th1 and Th2 T helper cell subsets affect patterns of injury and outcomes in glomerulonephritis.Kidney Int,1999,55(4):1198
, 百拇医药
11 Wallis WJ,Furst DE,Strand V et al.Biologic agents and immunotherapy in rheumatoid arthritis:Progress and Perspective.Rheum Dis Clin North Am,1998,24(3):537
12 Tipping PG,Kitching AR,Huang XR et al.Interleukin-4 and interleukin-10 attenuate established crescentic glomerulonephritis in mice.Kidney Int,1997,52:52
13 Tam FWK,Smith J,Karkar AM et al.Interleukin-4 ameliorates experimental glomerulonephritis and regulates glomerular gene expression of IL-1 decoy receptor.Kidney Int,1997;52:1224
, 百拇医药
14 Fouqueray B,Suberville S,Isaka Y et al.Reduction of proteinuria in anti-glomerular basement membrane nephritis by interleukin(IL)-10 gene transfer(abstract).J Am Soc Nephrol,1996,7:1698
15 Chadban SJ,Tesch GH,Lan HY et al.Effect of interleukin-10 treatment on crescentic glomerulonephritis in rats.Kidney Int,1997,51:1809
16 Ishida H,Muchamuel T,Sakaguchi S et al.Continuous administration of anti-interleukin-10 antibodies delays onset of autoimmunity in NZB/WF1 mice.J Exp Med,1994,179:305
17 Chu CQ,Londei M.Induction of Th2 cytokines and control of collagen-induced arthritis by nondepleting anti-CD4.J Immunol,1996,157:2685
(1999-05-15收稿), 百拇医药
单位:南京军区南京总医院 解放军肾脏病研究所(南京,210002)
关键词:Th1/Th2;肾脏病;治疗
肾脏病与透析肾移植杂志990329
自从1986年Mosmman和Coffman依据分泌细胞因子的差别将小鼠CD4+T细胞进一步分成Th1(主要产生IL-2及IFN-γ)及Th2(主要产生IL-4及ΙL-5)细胞亚群以来[1],Th1和Th2细胞效应及其平衡,在实验动物模型及人类感染和自身免疫性疾病中得到了深入研究。研究肾脏疾病中Th1与Th2平衡的意义,不仅在于更深入地理解疾病免疫发病机制,更重要的在于探索肾脏疾病治疗新途径[2]。那么,调整Th1/Th2平衡能否用于肾脏疾病的临床治疗呢?
, http://www.100md.com
1 Th1/Th2细胞效应的特征及其平衡
Th1细胞效应所产生的细胞因子除了IL-2与IFN-γ外,还包括TNF-β和IL-17(后者也是一种促炎症因子,能增加IL-6,GM-CSF及PGE2的产生)。Th2细胞效应所产生的细胞因子除了IL-4及IL-5外,还包括IL-6,IL-10及IL-13。Th1细胞效应主要是诱导巨噬细胞及细胞毒性T淋巴细胞活化,B细胞所产生的IgG转向(swith)易于补体活化及发挥调理作用的IgG亚类(小鼠的IgG2a和IgG3,人类的IgG1和IgG3),因此Th1细胞效应主要促进细胞内致病原的清除及器官特异性自身免疫反应(如迟发型变态反应DTH);而Th2细胞效应所产生的细胞因子(IL-4,IL-5,IL-10)参与了过敏性变态反应及寄生虫感染性疾病,促进B细胞产生IgE及IgG1(小鼠)或IgG4(人类)[3]。Th1和Th2的前体细胞(Tho)本身也是由原始T细胞活化而来,本身又能产生IL-4、IL-2和IFN-γ,原始T细胞活化后能产生IL-2。最近,Th3细胞也为人们所认识,它所产生的主要细胞因子为TGF-β[4]。
, http://www.100md.com
Th1/Th2细胞效应的特征之一是自身促进作用,即各亚群Th细胞因子诱导Th0细胞向各自的细胞亚群分化并促进各自Th细胞亚群活化。IFN-γ诱导Th0细胞向Th1细胞亚群分化,IL-4诱导Th0细胞向Th2细胞亚群分化[5]。
Th1/Th2细胞效应的特征之二是相互抑制作用,即各类Th细胞因子对另一类细胞亚群的活化及细胞因子的产生具有抑制作用。IL-4及IL-10都是IFN-γ的强力抑制剂,而IFN-γ亦反过来能抑制IL-10的产生[6]。
Th1/Th2细胞效应在通常情况下保持着相对的平衡(附图)[7]。这一平衡的维持是相当错综复杂的,就是在疾病状态下,Th1/Th2功能的消长也不完全取决于Th细胞本身的功能及所产生的细胞因子。对这一平衡的研究表明,虽然启动Th1/Th2细胞效应抗原各不相同,但是,通过改变细胞因子环境及采用丝裂原刺激,在体内或体外即能改变Th0细胞的分化方向。在丝裂原刺激下,胎儿脐血T细胞如培养在IL-12及抗IL-4环境中则向Th1细胞分化;而培养在IL-4及抗IL-12的环境中则向Th2细胞分化,这提示调整Th1/Th2细胞效应平衡是可能的,而且可以绕过特异性抗原来调整这一平衡[8]。
, http://www.100md.com
附图 Th1/Th2细胞效应与细胞因子间的平衡及其相互作用
实线为促进或正反馈作用,虚线为抑制或负反馈作用
2 Th1/Th2细胞效应的调控途径
目前人们已认识到Th1/Th2细胞效应主要受以下环节的影响。
2.1 基因背景 Th1/Th2细胞效应的平衡受到诸多基因的影响。比如,在Th1细胞效应中,早期巨噬细胞分泌的IL-12是重要的决定因子,若在免疫反应启动初期缺乏抗原呈递细胞(APC)分泌的IL-12的作用,则T细胞可能因表达的IL-12受体中缺失β2亚单位而不再对IL-12发生进一步的反应。这种不反应性与11号染色体的一个区域(在人类为5p31区)有关联。实验性自身免疫性脑脊髓炎和非肥胖1型糖尿病的易感性也与此区域有关联。
2.2 抗原呈递 抗原的呈递(包括呈递的类型、剂量和途径)与Th1/Th2细胞效应有重要关系。采用极低剂量的抗原于粘膜或腹腔内刺激,可以诱导Th2细胞效应以产生抗炎细胞因子。如在类风显性关节炎(RA)患者采用小剂量胶原口服,可能诱导免疫耐受而治疗RA[9,11]。
, http://www.100md.com
2.3 细胞因子环境 如上所述,细胞因子环境对于两类细胞的分化具有肯定的影响,目前可以通过增强内源性细胞因子的表达及提供外源性细胞因子,或(和)采用相应细胞因子的拮抗因子,以改变细胞因子环境而调整Th1/Th2平衡。
2.4 细胞活化协同刺激分子 抗原呈递细胞表面表达的协同刺激分子B7-1(CD80)及B7-2(CD86)是两类Th细胞分化过程中的第二信使,两信使分子的T细胞表面配体为CTLA-4及CD28。小鼠体内研究证实,激活或阻断B7-1途径,可促进或抑制Th1细胞效应,而激活或阻断B7-2途径,可促进或抑制Th2细胞效应。
2.5 细胞活化信号通路 IL-12的作用需要经过Stat4基因的活化与表达,小鼠Stat4基因的不活化可导致IL-12产生障碍,最后引起Th1细胞效应障碍,而Stat6基因敲除小鼠IL-4的产生及Th2细胞效应亦出现明显障碍。IL-12受体中β2亚单位的表达是Th1细胞效应中对IL-12反应性的关键,而调节IL-12R β2可以控制以Th1细胞效应为主导的自身免疫性疾病的发生。
, 百拇医药
3 Th1及Th2平衡与肾脏疾病及其它自身免疫性疾病
当前判断疾病免疫发病机制中Th1或Th2细胞效应为主导的方法,主要是依据炎症组织局部细胞因子的表达,也可根据外周血清中抗体反应,如IgE的水平及IgG亚类的变化进行判断。在动物模型的研究中,根据干扰Th1或Th2细胞效应的各种环节后Th1或Th2细胞效应的变化来进行判断(见附表)。
附表 与Th1或Th2平衡相关的人类感染疾病及自免疫性疾病[9~11] Th1主导
Th1/Th2
Th2主导
人类肾脏病
伴新月体形成的弥漫增生性肾炎
, 百拇医药 IgA肾炎
膜性肾病
抗GBM疾病
狼疮性肾炎
微小病变
寡免疫复合物型新月体肾炎
Ⅴ型狼疮性肾炎
膜增殖性肾炎
类风湿性关节炎相关性肾炎
IFN-γ治疗后伴新月体形成的狼疮性肾炎
IL-2治疗后伴新月体形成的IgA肾炎
肾炎动物模型
, 百拇医药
异质期抗GBM肾炎的C57BL/6小鼠
抗GBM肾炎BALB/c小鼠(较少有新月体形成)
巨噬细胞依赖性抗GBM肾炎的Wistar-Kyoto大鼠
抗GBM肾炎的SD大鼠
狼疮性肾炎鼠模型
氯化汞诱发的肾小球肾炎鼠或兔模型
慢性GVH疾病
伴有膜性肾病改变的慢性HVG疾病
Hymann肾炎
其它疾病或情形
, 百拇医药 多发性硬化
哮喘
自身免疫性甲状腺炎
过敏性皮炎
重症眼肌无力
季节性结膜炎
类风湿性关节炎
正常妊娠免疫耐受
Lyme关节炎
特发性嗜酸性细胞增多症
反应性关节炎
SLE
, 百拇医药 接触性皮炎
硬皮病
1型糖尿病
慢性GVH病
结节性红斑
Omenn's Syndrome
习惯性流产
GVH:宿主抗移植物,HVG:移植抗宿主
4 肾脏疾病中针对Th1及Th2平衡的治疗措施
在Th1主导的实验性自身免疫性脑脊髓炎,自身免疫性甲状腺炎,1型糖尿病非肥胖小鼠模型及实验性结肠炎动物模型中,采用外源性IL-4或(和)IL-10治疗,能够选择性地抑制Th1细胞效应而减轻疾病的严重程度。这类研究的结果,为调整Th1/Th2细胞效应平衡提供了实验基础[9]。
, 百拇医药
4.1 直接针对Th1或Th2细胞功能与分化的措施 由于目前对于Th1及Th2细胞表面标记分子(如CD27及CD30)研究中存在有较多争议,因而直接作用于细胞表面分子的单克隆抗体的措施尚无任何报道。但是,T细胞IL-12受体的β2亚单位的表达与Th1细胞效应有比较密切的关系,而GATA-3与Th2细胞效应有比较密切的关系,而在各类肾脏疾病中观察这两种分子的表达有利于判断Th1与Th2平衡,并为相应的干预措施提供监测依据。另外,体外应用细胞因子激活的Th1或Th2细胞,然后转输注体内,也可能成为直接干预Th1或Th2平衡的措施之一。这两种可能的治疗措施尚未在动物模型上应用。
4.2 针对细胞因子平衡的措施
4.2.1 直接针对细胞因子 针对细胞因子的措施包括单独或联合投用外源性细胞因子(如IL-4,IL-6及IL-10,TGF-β),或其拮抗剂(如抗IL-10及抗IL-4的中和性抗体),或采用抗炎因子基因疗法。其中最有潜力的是Th2细胞因子系列中的IL-4和IL-10,而IL-6及TGF-β治疗作用相对不确定而潜力较小。基因疗法尚在初步的探讨中。
, http://www.100md.com
Tipping等[12]在异质期抗GBM肾炎的C57BL/6小鼠模型上证实,联合应用IL-4及IL-10可减少新月体的形成及保护肾功能,其中IL-10具有独立的保护功能,而IL-4不具有独立的保护作用。这一保护作用很可能不完全是Th1向Th2转化的结果,而是IL-4及IL-10本身的抗炎作用。上述假设在巨噬细胞依赖性抗GBM肾炎的Wistar-Kyoto大鼠模型上得以验证,肾小球中巨噬细胞活化标记在IL-10治疗后也相应减少。
Tam等[13]在进展型抗GBM非新月体肾炎SD大鼠模型上证实,在抗原刺激之前采用IL-4治疗可以大幅度减少蛋白尿及肾脏病理损伤。这种作用与局部Ⅱ型IL-1结合性受体表达增多而结合IL-1,致使后者对浸润巨噬细胞的刺激减弱有关。
在Fouqueray[14]的研究中,应用人类IL-10基因转染(通过转染肾系膜细胞或全身骨骼肌细胞)至同质期抗GBM大鼠模型后,蛋白尿排泄减少。
, 百拇医药
但Chadban等[15]的研究指出,自注射抗GBM抗体后,每天投用重组鼠IL-10治疗SD大鼠模型并不能减少新月体的形成,相反,动物还会产生抗鼠IL-10的免疫反应,后者则可能加剧肾脏的损伤。
在多数抗GBM疾病的模型中,IL-4治疗的确具有肯定的抗炎作用,但其抗炎作用并不完全是通过作用Th1与Th2平衡发生的,还可能与抗炎脂质分子的产生有关。不过现在对于IL-4治疗的副作用知之甚少,IL-4能加强VCAM-1表达,从而加强T细胞及嗜酸性粒细胞与内皮细胞的粘附,IL-4还能刺激细胞外基质的产生,增加纤维母细胞产生 Ⅰ 型、Ⅲ 型胶原及纤维结合蛋白,这样可能引起T细胞在炎症局部的聚集和炎症局部组织的硬化。在Heymann肾炎中,增强IL-4 mRNA表达与肾小球中T细胞浸润无肯定的关系。
服用IL-10(而不是IL-4)对于大鼠抗GBM肾炎有保护作用,IL-10的保护作用更大程度上归因于它对Th1及Th2平衡的调整效应。因为在IL-10治疗后,从属Th1系列的免疫球蛋白亚类(IgG2a和IgG3)产生减少,而从属Th2系列的免疫球蛋白亚类(IgG1)的产生变化不明显。所以IL-10是目前所知的潜力最大的治疗制剂,已用于RA的Ⅱ期临床治疗[11]。不过,长期应用IL-10可引起Th2主导的肾小球肾炎的发作,如动物模型(NZB/WF1小鼠)治疗后发生狼疮样肾炎。而采用抗IL-10可延迟这类肾炎的发生。TGF-β与IL-10的抗炎作用相似。在实验性系膜增殖性肾炎模型中[16],TGF-β1的基因治疗可以在肾小球组织中表达足够活性的TGF-β,从而抑制肾小球细胞的增殖反应,这一作用可能与巨噬细胞分泌的炎症因子减少有关。
, http://www.100md.com
多数IgA肾炎患者存在Th1及Th2细胞效应的证据,但高血清IgA(特别是大分子IgA)患者的Th2效应占主导。IL-4不仅参与了免疫球蛋白的转向作用,而且还发挥了旁分泌与自分泌的作用,参与了IgA的沉积过程。而在伴新月体形成的IgA肾炎中,IgG1、IgG3沉积突出,且IFN-γ表达增强(Th1效应)。但目前对IgA肾炎尚无干扰尝试的报道。
在SLE中,单纯Th1或Th2效应的证据是难以确立的,但对不同类型狼疮性肾炎,Th1或Th2效应的主次仍有差别。在动物模型中,无论是单独增强Th1还是Th2细胞因子的作用,都可能加重病情,服用IFN-γ或IL-12也不能减少自身抗体的产生,但无论是阻断Th1还是Th2细胞因子,都可以减轻病情[10]。
在氯化汞诱导的肾小球肾炎动物模型中,Th2效应占主导地位。应用IFN-γ治疗这种肾炎小鼠虽然能减少IgE的产生,但不能减少自身抗体的产生及阻止肾炎的形成。
, http://www.100md.com
肾脏固有细胞虽然也能产生部分Th1与Th2系列的细胞因子,但是现在尚没有证据表明致肾炎性免疫反应的发生与肾脏固有细胞产生的细胞因子有关。在炎症过程中,肾脏固有细胞与免疫细胞之间的确有相互作用,但目前更多的人认为是全身性免疫反应决定了Th1与Th2效应。
4.2.2 诱导体内保护性细胞因子 尽管采用外源性针对细胞因子的措施可以取得肯定的作用,但是对慢性自身免疫性疾病来说,长期体外服用细胞因子是不现实的,而且抗原长期刺激的体外研究已表明维持长期Th1与Th2间的转化是不可能的。另外,疾病早期与后期的Th1/Th2平衡可能不尽相似。IL-12在反复发作的感染性疾病及自身免疫性疾病的复发过程中起着重要的作用,而应用抗IL-12在Th1主导的疾病发生早期或复发前,可能阻止其发作或复发,而疾病发生后应用抗IL-12则可能加重Th1主导的疾病的程度。
4.3 诱导旁观抑制效应 激活的Th2/Th3细胞对于Th1细胞依赖的炎症反应具有保护应用,这一作用的发生主要是通过抗原刺激下的IL-4,IL-10及TGF-β而发挥的。如在粘膜表面采用一定剂量的抗原摄入可诱导Th3细胞的产生,这一作用不依赖于IL-4,而依赖于TGF-β,而采用修饰抗原刺激则出现依赖于IL-4的途径。不过,这样激活的T细胞主要位于粘膜局部,而使这类细胞迁移至炎症局部还有没能解决的困难。有人发现在早期的抗原接触过程中,应用非耗竭性抗CD4多克隆抗体可以使Th1细胞效应转化为产生IL-4的Th2细胞效应[17]。
, 百拇医药
总之,对于Th1/Th2平衡与肾脏疾关系的研究正在不断深入,而对于这一平衡的治疗措施,尚处于动物实验的初级阶段。就是动物模型上,这些措施多数仅在疾病发作的早期有治疗效应。应用于临床肾脏疾病的治疗仍有不少尚未克服的问题。不过,这一领域的研究进展日新月异,部分细胞因子生物制剂已用于RA的Ⅱ期临床研究。调整Th1/Th2效应平衡在肾脏病治疗中的应用指日可待。
参考文献
1 Mosmann TR,Cherwinski H,Bond MW et al.Two type of murine helper T cell clone.I.Definition according to profies of activities and secreted proteins.J Imunol,1986,136:2348
2 Baud L,Fouqueray B,Bellocq A.Switching off renal inflammation by anti-inflammatory mediators:the facts,the promise and the hope.Kidney Int,1998,53:1118
, 百拇医药
3 Romagnani S.Human Th1 and Th2 subsets:doubt no more.Immunol Today,1991,12:256
4 Abbas AK,Murphy KM,Sher A.Functional diversity of helper T lymphocytes.Nature,1996,383:787
5 Daynes RA,Araneo BA,Dowell TA et al.Regulation of murine lymphokine production in vivo.Ⅲ.The lymphoid tissue microenvironment exerts regulatory influences over T helper cell function.J Exp Med,1990,171:979
6 Chomarat P,Issoan MC,Banchereau J et al.Interferon γ inhibits interleukin-10 production by monocytes.J Exp Med,1993,177;523
, http://www.100md.com
7 刘俊铎,刘志红.Th1/Th2细胞与自身免疫性疾病.肾脏病与透析肾移植杂志,1997,6(6):555
8 Sszabo SJ,Dighe AS,Gubler U et al.Regulation of the interleukin-12R β 2subunit expression in developing T helper 1 and Th2 cells.J Exp Med,1997,185:817
9 Miossec P,van den Berg W.Th1/Th2 cytokine balance in arthritis.Arthritis Rheum,1997,40(12):2105
10 Holdsworth SR,Kitching R,Tipping PG.Th1 and Th2 T helper cell subsets affect patterns of injury and outcomes in glomerulonephritis.Kidney Int,1999,55(4):1198
, 百拇医药
11 Wallis WJ,Furst DE,Strand V et al.Biologic agents and immunotherapy in rheumatoid arthritis:Progress and Perspective.Rheum Dis Clin North Am,1998,24(3):537
12 Tipping PG,Kitching AR,Huang XR et al.Interleukin-4 and interleukin-10 attenuate established crescentic glomerulonephritis in mice.Kidney Int,1997,52:52
13 Tam FWK,Smith J,Karkar AM et al.Interleukin-4 ameliorates experimental glomerulonephritis and regulates glomerular gene expression of IL-1 decoy receptor.Kidney Int,1997;52:1224
, 百拇医药
14 Fouqueray B,Suberville S,Isaka Y et al.Reduction of proteinuria in anti-glomerular basement membrane nephritis by interleukin(IL)-10 gene transfer(abstract).J Am Soc Nephrol,1996,7:1698
15 Chadban SJ,Tesch GH,Lan HY et al.Effect of interleukin-10 treatment on crescentic glomerulonephritis in rats.Kidney Int,1997,51:1809
16 Ishida H,Muchamuel T,Sakaguchi S et al.Continuous administration of anti-interleukin-10 antibodies delays onset of autoimmunity in NZB/WF1 mice.J Exp Med,1994,179:305
17 Chu CQ,Londei M.Induction of Th2 cytokines and control of collagen-induced arthritis by nondepleting anti-CD4.J Immunol,1996,157:2685
(1999-05-15收稿), 百拇医药